Природа изменчивости, направления и формы ее познания
Концепция гена, учение о материальных основах наследственности и их методологическая интерпретация неотделимы от ответа на вопрос, как возникают и реализуются в индивидуальном и историческом развитии изменения наследственности, каковы их причины и сущность. Генетика постоянно связывала решение этого вопроса с изучением механизмов передачи наследственных признаков, наследственного воспроизведения живых систем.
Изменчивость - одно из характернейших свойств организмов, один из фундаментальных факторов их эволюции и селекции и вместе с тем результат последних, замечательное приспособление живых систем, условие сохранения динамического равновесия стационарных процессов жизнедеятельности во взаимодействии с факторами внешней среды. Жизнь сохраняется не только за счет саморегулирования живых систем, их самовоспроизведения, но и с помощью как обратимых, так и необратимых, прогрессирующих изменений, осуществляющихся в форме перестройки их наследственной основы под влиянием целого комплекса внутренних и внешних факторов.
Научное доказательство значения изменчивости в индивидуальном и историческом развитии организмов было впервые Сделано, как известно, в теории Ч. Дарвина об эволюции путем естественного отбора. В противовес ламаркистской интерпретации природы и характера изменчивости, которая - по крайней мере, в своей материалистической форме - представляла организм как пассивный продукт однозначных наследственно-приспособительных реакций на внешние воздействия, отождествляла индивидуальную и видовую, историческую изменчивость, Дарвин построил свою теорию на новом представлении об изменчивости, учитывающем сложные диалектические связи и взаимодействия организма со средой.
Это представление явилось не только научной основой подлинной теории эволюции, но и послужило исходной базой для детального исследования механизмов наследственности на новом этапе биологического познания, в частности в рамках генетики. Современная генетика в значительной мере исходит из дарвиновского i представления, поэтому отметим его основные принципы, играющие и сегодня определяющую роль в методологической интерпретации проблемы изменчивости организмов.
Дарвин различал определенную и неопределенную изменчивость, характер которых он связывал с разной формой взаимодействия организма и среды. В определенной изменчивости различия, вызываемые изменением факторов внешней среды, имеют, по Дарвину, относительно однотипный для группы организмов, общий для того или иного вида характер. Здесь на первый план выдвигаются особенности воздействующих условий, а индивидуальная специфичность наследственной природы организмов оказывается относительно несущественной, не главной для результатов в целом. В отличие от этого характер неопределенной изменчивости обусловливается как раз в основном внутренней спецификой организма. Эта индивидуальная, единичная изменчивость лишь косвенно связана с изменениями внешней среды. Она, как правило, является наследственной и создает различия, обеспечивающие базу для действия естественного отбора и эволюции в направлении приспособления. Последнее означает, что, хотя Дарвин четко отличал индивидуальную изменчивость от видовой, эволюционной, он не проводил вместе с тем непереходимой черты между ними, не отрывал их друг от друга и тем более не противопоставлял одну форму изменчивости другой.
Дарвиновское учение об изменчивости создало научную основу для детального изучения причин и характера этого фундаментального свойства организмов. Само по себе, однако, оно не раскрывало механизма индивидуальной изменчивости, предполагая ее в теории естественного отбора как исходную предпосылку, взятую из многочисленных наблюдений. Эта черта классического дарвинизма была отмечена Энгельсом, который считал, что, "когда Дарвин говорит о естественном отборе, он отвлекается от тех причин, которые вызвали изменения в отдельных особях, и трактует прежде всего о том, каким образом подобные индивидуальные отклонения мало-помалу становятся признаками определенной расы, разновидности или вида. Для Дарвина дело идет прежде всего не столько о том, чтобы найти эти причины,- они до сих пор частью совсем неизвестны, частью же могут быть указаны лишь в самых общих чертах,- сколько о том, чтобы найти ту рациональную форму, в которой их результаты закрепляются, приобретают прочное значение"*. В полном соответствии с этим высказывался также К. А. Тимирязев, который считал, что "дальнейшее развитие биологии должно заключаться в более глубоком анализе двух основных свойств организмов, от которых дарвинизм отправляется как от данных: в анализе изменчивости и наследственности - и прежде всего первой, как самого первичного фактора эволюции"**.
* (Маркс К., Энгельс Ф. Соч. 2-е изд. Т. 20. С. 70.)
** (Тимирязев К. А. Соч. М., 1939. Т. IX. С. 110.)
Генетика и пошла по этому пути детального анализа причин и характера изменчивости. Разные направления и формы принимал он на протяжении ее истории, причем, как это уже отмечалось выше, генетический анализ в определенный период развития теории наследственности и изменчивости неоправданно отрывался от своей эволюционной основы, противопоставлялся дарвинизму.
Исследование деталей, механизмов изменчивости сопровождалось во многих случаях утратой целостных подходов, столь ярко проявившихся в дарвиновском учении.
В предшествующем изложении было отмечено, однако, не только это, но и то, как постепенно преодолевались указанные ограниченности теории генетики по мере успехов в познании наследственности и изменчивости на разных уровнях их проявления в жизнедеятельности организмов и популяций. Соответственно этим успехам познания и в тесной связи с эволюцией понятия гена, учения о материальных основах наследственности исторически изменялись и представления о природе мутации, которыми вслед за де Фризом стали называть всякие наследственные изменения ("неопределенная изменчивость", по терминологии Дарвина), отличая их от ненаследственных модификаций, в известной мере соответствующих дарвиновской "определенной изменчивости".
Не повторяя того, что было сказано при рассмотрении исторического развития методологических основ генетики, обратим внимание лишь на один существенно важный для современной трактовки понятия мутаций момент, связанный с эволюцией самого этого понятия. Первоначально, когда генетический анализ принимал зачастую антидарвинистскую направленность, центральное место в нем занимали те наследственные изменения, которые не имеют характера мелких индивидуальных отклонений и вместе с тем не связаны непосредственно, прямо с изменениями внешних факторов. В частности, де Фриз и называл мутациями только такие крупные изменения наследственности. Он полагал, что мутации - это внезапные, скачкообразные изменения, которые возникают независимо от влияния среды и сразу оказываются наследственно устойчивыми. Они резко отличаются от так называемых флюктуации, которые вызываются влиянием внешних факторов, имеют характер небольших отклонений от среднего выражения того или иного признака и не передаются по наследству. Мутации де Фриз связывал со "спонтанными" изменениями единичных "пангенов", которые в ядре клетки находятся, как он полагал, в состоянии покоя, а в протоплазме - периодически изменяются.
Такая трактовка природы и значения мутаций, их противопоставление мелким изменениям приводили к отрицанию творческой роли естественного отбора, к его уподоблению, как говорил де Фриз, "большому ситу", отсеивающему уже готовые формы, появившиеся в результате мутаций. Тем самым мутационизм выступил первоначально прямым антиподом дарвинизма, учения об эволюции путем естественного отбора. В таком виде он был принят рядом ученых на первых этапах развития генетики, причем, как отмечалось, представление о неизменности признаков между двумя мутациями было перенесено на наследственный фактор. Мутации связывались уже не с изменениями единичных структур наследственной основы (генов), но с их перегруппировкой в хромосомах.
Однако наряду и в противовес этому представлению в генетике складывалось и иное понимание природы и значения мутаций, которое получало все более конкретную цитологическую интерпретацию и отходило как от переоценки роли отдельных крупных мутаций и их противопоставления мелким наследственным изменениям, естественному отбору, так и от абсолютизации устойчивости, стабильности отдельных генов. На этом этапе понятие мутаций получило более разностороннюю и углубленную трактовку. Оно стало включать в себя уже не только крупные изменения наследственности, но и мелкие, роль которых в эволюции путем отбора является основной, главной. Оно стало прочно опираться также на представление не только о хромосомных перестройках, но и о качественных изменениях отдельных генов.
Это связывало воедино концепцию мутаций с теорией гена и эволюционным учением. В частности, Г. Мёллер, который был одним из пионеров такого рода исследований природы мутаций, прямо ставил вопрос о принципах, управляющих эволюцией, в зависимость от представления о качественных изменениях генов как необходимого условия эволюции. Он видел в изучении причин первичных, генных мутаций одно из решающих направлений исследования наследственности и эволюции. "Мы не можем,- писал Мёллер,- оставить эти невидимо-маленькие и все же имеющие фундаментальное значение частицы - гены - навеки нетронутыми в их недоступных убежищах, ибо мириады этих генов, располагающихся в тонкие цепочки, непрерывно излучают те действенные, упорядоченные, но ускользающие от нас силы, которые строят и разрушают наш живой мир"*.
* (Мёллер Герман Джозеф. Избранные работы по генетике. М.; Л., 1937. С. 175.)
Такую же высокую научную оценку исследованиям генных мутаций давал и Н. К. Кольцов, видевший в них могучее средство для изменения наследственной природы организмов и решающий путь экспериментального анализа проблем эволюции. Вообще, можно сказать, подавляющая часть ученых, занимавших передовые позиции в генетике, сразу же увидела здесь начало нового этапа развития познания природы мутаций.
Ясно осознавалось, однако, что "это - трудный путь, но,- как предсказывал Г. Мёллер,- с помощью волшебства науки он должен быть преодолен"*. Современная генетика близка к этому, хотя ей пришлось преодолевать не только то, что отчетливо виделось в качестве препятствий в самом начале нового пути, но и то, что возникло уже в процессе продвижения по нему, а многое еще сегодня не поддается "волшебству науки".
* (Мёллер Герман Джозеф. Избранные работы по генетике. М.; Л., 1937. С. 175.)
Современная генетика исследует природу мутаций в разных направлениях и разных формах, которые в своем единстве создают отчетливую - по крайней мере в основных контурах - картину того, что уже достигнуто в исследовании сущности и причин наследственных изменений и что еще необходимо детально исследовать и по каким эвристически эффективным методологическим линиям следует вести это исследование. Разумеется, в последнем случае какой-либо однозначности в предсказаниях нет, и, может быть, ее и не следует пока добиваться, так как еще много неожиданного, непредвиденного откроется перед взором генетиков в исследовании проблемы мутаций, интенсивно развивающихся всего несколько последних десятилетий.
Прежде чем перейти к рассмотрению методологической интерпретации проблемы мутаций и их эволюционного значения, отметим некоторые, наиболее существенные моменты той целостной картины наследственных изменений, которая уже создана современной генетикой.
Согласно современным экспериментальным данным, полученным в ходе анализа молекулярных взаимодействий живых систем, мутации, как известно, тесно связываются с химическими изменениями генетического материала. Соответственно различаются хромосомные и генные (точечные) мутации. Первые затрагивают всю хромосому или ее большую часть; вторые - небольшую часть хромосомы - ген, т. е. определенный локус ДНК.
Хромосомные мутации сопровождаются изменениями видимой структуры одной хромосомы или их полного набора в клетке (изменения кариотипа, числа хромосом в ядре, приводящие, в частности, к полиплоидии, т. е. увеличению в два, три и более раз гаплоидных наборов хромосом). Изменения структуры одной хромосомы (хромосомные аберрации) связаны с нарушениями линейного порядка генов. Эти внутрихромосомные перестройки могут происходить в форме разрыва хромосом за счет выпадения какого-то участка хромосом (делеция) или его добавления (дупликация). Наблюдаются и такие внутрихромосомные перестройки, при которых тот или иной участок хромосомы либо меняет свою ориентацию, разворачивается вокруг своей оси на 180° (инверсия), либо он меняется местами с другим участком хромосомы (транслокация).
Схематически различные типы хромосомных мутаций иллюстрируются следующим образом*:
* (См.: Вилли К. Биология. М., 1966. С. 519.)
a b c d e f Норма
a b d e f Делеция (участка с)
a b с с d e f Дупликация (участка с)
а е d с b f Инверсия (участка b-e)
Механизм генных мутаций, при которых изменение признака возникает за счет изменения внутри самого гена, детерминирующего этот признак, заключается в изменении порядка оснований в структуре ДНК, в нарушении строгой комплементарности двух пар оснований (аденина - тимина и гуанина - цитозина), обеспечивающей репликацию ДНК при удвоении хромосомы в митозе.
В результате изменения последовательности пар оснований возникает ситуация, при которой либо вообще оказывается невозможным синтез белка, либо происходит формирование необычного, измененного белка, что и обусловливает появление новых признаков.
На рисунке 1 наглядно представлен молекулярный механизм этого процесса. На нем показано, что какой-либо аналог пурина или пиримидина может нарушить процесс репликации и вызвать мутацию (изменение последовательности нуклеотидов в ДНК показано черным).
Генные мутации в естественных условиях наблюдаются чрезвычайно редко (их частота современными методами определяется как одна мутация на 100 тыс.- 1 млн половых клеток в каждом поколении). Однако частота генных мутаций интенсифицируется за счёт повышения температуры, под воздействием ионизирующей радиации или химических мутагенов. Искусственно индуцированные генные мутации интенсивно исследуются в настоящее время как сложный процесс, имеющий несколько этапов, на каждом из которых проявляется специфичность того или иного мутагена. При этом учитывается не только механизм первичной мутации (реакция мутагена с ДНК), но и ее последующая реализация с учетом внутриклеточных взаимодействий биохимических и физиологических процессов, надмолекулярных и биологических структур и т. д., действия открытых в последнее время особого класса химических веществ (антимутагенов), которые, проникая в клетку или присутствуя в ней постоянно, погашают процесс мутагенеза.
Поэтому первичные мутации, изменения взаимоположений азотистых оснований в молекуле ДНК, интерпретируются сейчас лишь как начало, предпосылка, основа мутационного процесса в целом, который отнюдь не сводится к химии молекул ДНК. Подчеркивая исключительное значение химии ДНК для проблемы мутаций, Н. П. Дубинин отмечает, что, поскольку мутационный процесс протекает в живой клетке, "на его специфику существенный отпечаток накладывает эволюция каждого из видов. Биологическая сторона в мутационном процессе исключительно велика. Сами факторы естественного мутирования во многом черпаются внутри самой клетки в виде появления в ней метаболитов, имеющих мутагенное значение. Количество мутаций и их качественная характеристика оказываются специфичными для различных видов и их отдельных популяций. Мутационный процесс, протекая на молекулярном уровне, вместе с тем осуществляется как биологическое явление. Биология естественного мутационного процесса обусловлена тем, что его причины коренятся в биохимическом взаимодействии ядра, цитоплазмы и внешней среды и что факторы, ведущие к молекулярным изменениям в хромосомах, воспринимаются не просто молекулами ДНК, непосредственно, а через надмолекулярную биологическую организацию хромосом"*.
* (Дубинин Н. П. Молекулярная генетика и действие излучений на наследственность. М., 1963. С. 194. См. его же: Общая генетика. М., 1968.)
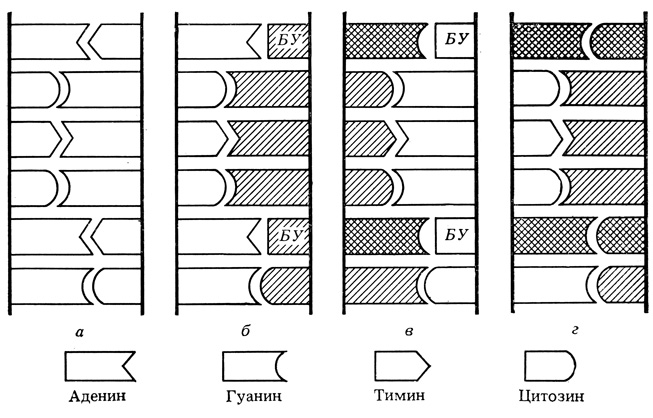
Рис. 1. а - исходная ДНК; б - бромурацил включился в новую цепочку вместо тимина; в - цепочка с бромурацилом ведет к образованию новой цепочки (гуанин образует пару с бромурацилом); г - новая мутантная ДНК с измененной по сравнению с исходной последовательностью нуклеидов (ГЦ вместо AT)
Однако, констатирует Н. П. Дубинин, принципы и сущность конкретной организации хромосом как надмолекулярных и биологических структур еще неясны. Зная в грубом абрисе элементы системы целостной жизнедеятельности клетки, мы еще не знаем, как они детерминируют нарушения соответствия в синтезе белков и ДНК, выражающиеся в мутациях. "Молекулярная генетика,- пишет Н. П. Дубинин,- изучает в этой системе ключевые позиции, однако управлять жизнью - значит управлять системой клетки и организма в целом... Загадка развития особи, клеточная дифференцировка не могут быть раскрыты, пока мы не поймем системных изменений в клетках"*.
* (Дубинин Н. П. Молекулярная генетика и действие излучений на наследственность. М., 1963. С. 233. )
Современная теория устанавливает механизмы детерминаций-признаков молекулярными процессами. Достигнуты большие успехи и в объяснении внутримолекулярных процессов, обеспечивающих синтез белка, специфической роли в этих процессах не только ДНК, но и РНК, различных субклеточных структур и пр.
Однако то, как осуществляется перевод генетической информации с языка генов на язык признаков, во многом еще остается загадкой. В сущности, генетика пока что имеет дело здесь со своеобразным "черным ящиком", где более или менее известны явления на входе и выходе - первое и последнее звено причинной цепи - и остается неясным скрытый механизм системы*.
* (Примеч. 1988 г. Конечно, за истекшие годы здесь появилось очень много нового, но принципиально дело не изменилось.)
Но принципы генетики все более проясняются, хотя еще остается много проблем, по которым возможны разные точки зрения, а следовательно, необходимо обсуждение, в том числе и в методологических аспектах, различных подходов.
Предпосылкой такого обсуждения служат, однако, не только экспериментальные и теоретические данные, касающиеся количественных сторон, отдельных элементов генетических систем, принимающих участие в мутагенезе, и их связей, целостных взаимодействий. Большое значение имеет также качественная характеристика мутаций с учетом их роли в эволюционном процессе. Хотя в общем и целом современная генетика отказалась от переоценки значения крупных мутаций в возникновении новых видов, альтернативно противопоставляемых постепенному накапливанию и отбору мелких мутаций, все же ряд ученых полагает, что эволюция осуществляется в результате именно этих макромутаций генетической системы. Что же касается мелких мутаций, то им отводится главная роль лишь в возникновении разновидностей или географических рас. Эти позиции нашли свое отражение, в частности, в трудах Р. Гольдшмита, который, исследуя генетические принципы эволюции, развил ряд доводов в пользу ведущей роли макромутаций и против представления о накапливании и отборе микромутаций, получившего экспериментальное и математическое обоснование в генетике популяций, а с качественной стороны - в особенности в работах И. И. Шмальгаузена.
Одним из основных доводов против этого представления является ссылка на неблагоприятное влияние суммирования отдельных мутаций, которые, как правило, вредны. Однако, как подчеркивал И. И. Шмальгаузен, рассматривая вопрос об эволюционном значении мутаций, в природных условиях естественный отбор никогда не имеет дело с отдельными мутациями. "Если все мутации, взятые в отдельности,- писал он,- как правило, вредны, т. е. связаны с нарушением установившихся соотношений, то это ясно показывает, что ни одна мутация сама по себе не является этапом на пути эволюции. Процесс эволюции ни в коем случае нельзя себе представлять (как это делают защитники мутационной теории) результатом простого суммирования мутаций. Каждая мутация подлежит сначала известному преобразованию и комбинированию под руководящим влиянием естественного отбора"*.
* (Шмальгаузен И. И. Проблемы дарвинизма. М., 1946. С. 224.)
В качестве основной базы эволюционного процесса, естественно, не могут быть безусловно вредные мутации, которые поэтому сразу же устраняются. Но, отмечал И. И. Шмальгаузен, такую базу создают: 1) условно вредные мутации, т. е. неблагоприятные для организма в обычных, исторически сложившихся условиях существования и безразличнее или даже благоприятные в новых условиях; 2) частично вредные, т. е. неблагоприятные в некоторых своих выражениях и полезные - в других; 3) малые мутации, которые, подчеркивал И. И. Шмальгаузен, "вносят столь незначительные нарушения, что заметно не отзываются на жизнеспособности особей, могут сохраняться и накапливаться в популяции. При скрещивании они различным образом комбинируются и в результате могут получиться вполне гармоничные сочетания. Еще важнее то обстоятельство, что при комбинировании мутаций их выражение нередко существенно изменяется. Несомненно, что малые мутации имеют значение основного материала для эволюционного процесса"*; 4) комбинирование частично вредных мутаций с другими, безразличными или просто малыми мутациями, которое может привести к нейтрализации вредных выражений мутаций и появлению - в ходе естественного отбора - полезных изменений.
* (Шмальгаузен И. И. Проблемы дарвинизма. М., 1946. С. 224. 225.)
Таким образом, в качественных характеристиках и выводах И. И. Шмальгаузена основной акцент делался на значение малых мутаций и их комбинирование в процессе эволюции, где в результате естественного отбора уничтожаются вредные выражения мутаций и усиливаются полезные. В итоге, отмечал И. И. Шмальгаузен, "положительные мутации входят после этой переработки в состав нормы, в обезвреженные мутации накапливаются в популяции частью в скрытом виде и создают известный резерв внутривидовой изменчивости. При всяком изменении внешних условий этот резерв может быть быстро мобилизован"*.
* (Шмальгаузен И. И. Проблемы дарвинизма. М., 1946. С. 227.)
Эти выводы и характеристики основываются на данных количественного анализа мутаций и их эволюционного выражения, которые получены интенсивно развивающейся, в особенности в последние годы, генетикой популяций, исследующей генетические принципы эволюции. В ее рамках было показано, что в ходе мутационного процесса в природных популяциях накапливается своеобразный потенциал изменчивости ("скрытая изменчивость", по С. С. Четверикову), механизм формирования которого соответствует основным законам генетики. "Множество мутаций,- пишут Н. П. Дубинин и Я. Л. Глембоцкий,- как только что возникших (давление мутаций), так и поддерживаемых в популяции под действием отбора (давление отбора) или случайно размножившихся в популяциях (давление генетико-автоматических процессов), в сложном взаимодействии включено в наследственное многообразие любой популяции. Все эти мутации в своем наследовании подчинены законам Менделя, их соотношения и рекомбинации в единой популяции, связанной половым размножением, оказываются вполне закономерными, выражаясь в определенных генетических структурах популяций, разных при разном соотношении давления мутаций, отбора и случайного размножения разных генов"*.
* (Дубинин Н. П., Глембоцкий Я. Л. Генетика популяций и селекция. М., 1967. С. 23.)

Сергей Сергеевич Четвериков (1880-1959)
Соответственно с генетической точки зрения популяция предстает как размножающееся сообщество индивидов, имеющее практически общую генетическую основу и вместе с тем обладающее весьма значительным генетическим разнообразием. Поскольку, как доказал С. С. Четвериков, именно популяция, а не отдельная особь или вид оказывается элементарной эволюционирующей единицей, в качестве элементарного эволюционного явления должно рассматриваться устойчивое изменение генотипического состава популяций как целого*. Основой таких изменений служат микромутации, связанные с изменением признаков, определяемых многими генами, с процессами взаимодействия внутри генотипа.
* (См.: Тимофеев-Ресовский Н. В. Микроэволюция: Элементарные явления, материал и факторы микроэволюционного процесса // Ботанический журн. 1958. № 3. С. 318-321.)
Генетика популяций установила, что сочетание в едином генотипе разных аллельных генов повышает жизнеспособность организмов (гетерозис), что, следовательно, отдельные вредные в гомозиготном состоянии мутации могут оказаться полезными в гетерозиготном состоянии, причем их ценность определяется свойствами генофонда популяции в целом. Установление взаимовлияния между отдельными генами и генотипами, с одной стороны, и общим генофондом популяции - с другой,- важнейшее достижение генетики популяций, являющееся основой современного учения об эволюции путем естественного отбора.
Разумеется, к этому не сводятся генетические принципы эволюции, но изложить их здесь достаточно полно не представляется возможным*. Принципиально важно тем не менее иметь в виду для последующего методологического обсуждения проблемы мутаций в их связи с эволюцией то исключительно плодотворное обобщение, которое сделал в последние годы своей жизни И. И. Шмальгаузен, использовав для этой цели понятия и методы теории информации. Попытаемся хотя бы вкратце рассмотреть наиболее существенные элементы этого обобщения и ту принципиальную схему индивидуального цикла передачи наследственной информации и регулирующего механизма эволюции популяций, которая была предложена И. И. Шмальгаузеном.
* (Хотелось бы обратить внимание читателя на исключительно интересные работы по этому вопросу, выполненные в разное время А. А. Малиновским и обобщенные, в частности, в статье "Типы управляющих биологических систем и их приспособительное значение" (Проблемы кибернетики. М., 1960. Вып. 4.).)
Сейчас уже не вызывает сомнений тот факт, что изменение генетического материала осуществляется не самопроизвольно, а под воздействием среды на все стадии гаметогенеза и на сами гаметы, что вызывает нарушение в передаче наследственной информации и обусловливает появление мутаций. Последние, как уже отмечалось, в большинстве своем оказываются вредными в силу того, что они ведут к дезинтеграции живых систем, нарушению состояния динамического равновесия с окружающей средой. Это связано с тем, что, как пишут Р. Сэджер и Ф. Райн, "современный генотип, вероятно, очень близок к оптимальному для существующих условий. Ударяя молотом по швейцарским часам, мы имеем больше шансов ухудшить работу их механизма, нежели улучшить ее"*.
* (Сэджер Р., Райн Ф. Цитологические и химические основы наследственности. С. 52.)
Однако процесс мутирования может сопровождаться появлением благоприятных изменений, вероятность которых будет тем большей, чем выше частота мутаций. Именно они оказываются исходным материалом, обеспечивающим эволюцию, устойчивость биологической организации в условиях изменяющейся среды. Разрешение противоречия - необходимой устойчивости и одновременно мобильности генетического материала - достигается самой организацией наследственного кода и реализуется в популяции в ходе регуляции посредством циклического механизма с обратной связью, общая схема которого разработана И. И. Шмальгаузеном*.
* (См.: Шмальгаузен И. И. Перспективы применения точных методов для изучения факторов эволюции//Вестн. ЛГУ. Сер. биологическая. 1959. № 9. С. 110.)
Согласно этой схеме, прямая наследственная информация, передающаяся на молекулярном и внутриклеточном уровнях, осуществляется через зиготы. Она (в условиях непрерывного взаимодействия с факторами внешней среды и под контролем сложных регуляторных механизмов клетки и всего организма в целом) преобразуется в процессах индивидуального развития в фенотип особи, входящей в состав определенной популяции. Через посредство отдельных особей данной популяции осуществляется обратная информация, которая в биогеоценозе в результате естественного отбора вновь преобразуется в наследственную, причем в ходе преобразования осуществляется переход от уровня организации особи к внутриклеточному уровню. Эта преобразованная информация передается в популяцию в виде нового поколения измененных зигот, на чем и замыкается полный круг преобразований в элементарном цикле эволюции, идущей по пути адаптации и усложнения организации или стабилизации, селекции наиболее помехоустойчивых живых систем.
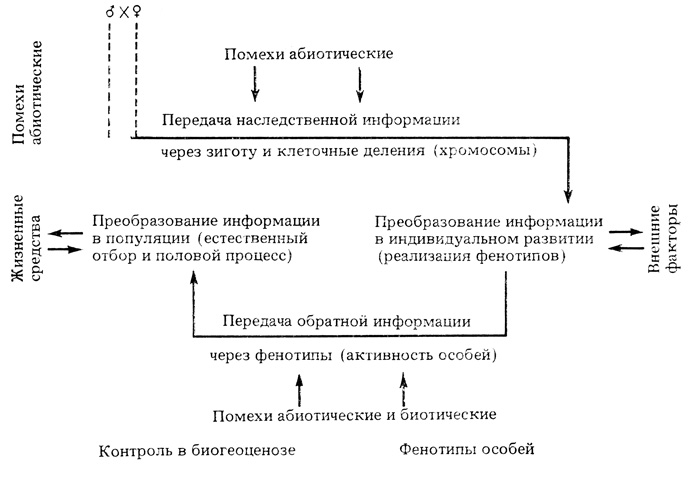
Общая схема регулирующего механизма эволюции популяций (по И. И. Шмальгаузену)
В комплексе взаимодействий, обеспечивающих индивидуальную и историческую изменчивость, огромную роль, по И. И. Шмальгаузену, играют не только внутренние, но и внешние факторы - как обычные, входящие в качестве необходимого элемента в преобразующие механизмы прямой (в индивидуальном развитии) и обратной (контроль в биогеоценозе) информации, так и необычные, приводящие к нарушению правильности работы регулирующих механизмов и их прогрессивному изменению, осуществляющемуся в форме приспособительных реакций на воздействие внешних факторов.
Этот последний вопрос прямо смыкается с целой серией взаимосвязанных с ним методологических проблем, касающихся теоретической интерпретации характера наследственной изменчивости (мутаций), прежде всего с проблемой детерминизма в генетике, необходимости и случайности в явлениях изменчивости, динамических и статистических закономерностей и т. д. Немало споров разгоралось по этим проблемам в ходе исторического развития генетики. И нельзя сказать, что они относятся лишь к прошлому науки, так как сами по себе эти проблемы еще далеки от своего окончательного разрешения. Можно поэтому определить общие, принципиальные подходы, которые дает диалектико-материалистическая методология для теоретической интерпретации сущности изменчивости в связи ее индивидуального и исторического выражения.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'