2.4. Уникальные гены и генные семейства
Итак, из предыдущих разделов явствует, что в состав типичного гена входят область транскрипции, состоящая из экзонов, интронов и зоны терминации транскрипции, а также различные регуляторные последовательности (энхансеры, сайленсеры и др.), которые чаще всего концентрируются недалеко от точки старта транскрипции, но могут располагаться и в области транскрипции, и на больших расстояниях перед нею (дальние энхансеры), и после нее (см. рис. 10). Поэтому, хотя область транскрипции, особенно если считать ее от старта до сайта полиаденилирования, легко ограничить, то всю область гена со всеми контрольными областями определить много труднее. Наиболее убедительным тестом являются опыты с трансгенными животными. Если работа пересаженного гена абсолютно не отличается от работы нормального гена, можно думать, что все его контрольные элементы вошли в состав пересаженного фрагмента ДНК-
Даже такой "полный" ген, как правило, не соседствует непосредственно со следующим геном, но погружен в последовательности ДНК, не выполняющие каких-либо очевидных функций, во всяком случае кодирующих. Гены в геноме, таким образом, напоминают острова архипелага в море неактивных, по крайней мере на первый взгляд, последовательностей.
Иногда гены являются действительно уникальными, т. е. представленными всего одной копией на гаплоидный набор ДНК. Однако это не самый частый случай. Чаще, особенно для высших эукариот, гены представлены не одиночными копиями, но семействами нескольких, сходных между собою генов, выполняющих сходные функции, т. е. кодирующих родственные друг другу белки.
Есть разные типы семейств генов. Одни из них - тандемно повторенные гены. В этом случае все представители семейства одинаковы по структуре и кодируют одни и те же продукты. Сюда относятся гены для рибосомной РНК, 5S РНК, гены для гистонов и некоторые другие. Так, например, в случае гистоновых генов в определенном участке хромосомы многократно (десятки или сотни раз в зависимости от вида) повторяется одинаковый блок, состоящий из пяти генов, разделенных спейсерами, содержащими контрольные элементы. Повторяющиеся единицы следуют непосредственно одна за другой без каких-либо вкраплений другой ДНК. Так устроены гены, продукты которых должны нарабатываться в больших количествах в короткие промежутки времени (рис. 11).
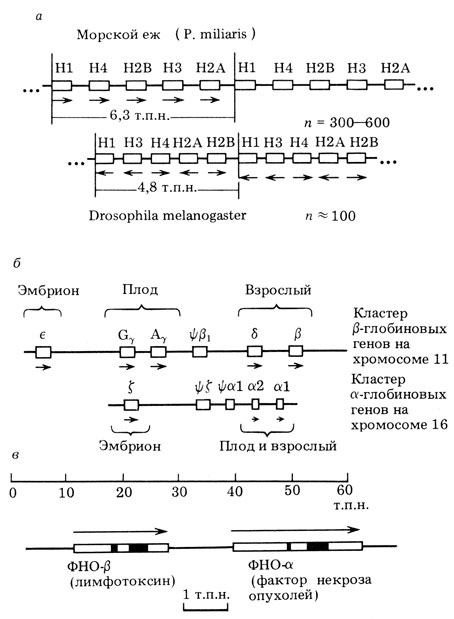
Рис. 11. Примеры генных семейств. а - гистоновые гены морского ежа и дрозофилы. Гистоновые гены - пример кластера многократно повторяющихся (л.) одинаковых генетических единиц; б - β-глобиновый и α-глобиновый кластеры человека, расположенные на двух разных хромосомах. В них наряду с активными генами присутствуют псевдогены (ty) ? Разные гены активны на разных этапах онтогенеза; в - небольшой кластер из двух генов фактора некроза опухолей (ФНО) человека. Гены выделены прямоугольниками (в случае ФНО указаны интроны - черные прямоугольники). Стрелки указывают направление транскрипции (по результатам, полученным М. Бирнстилом, Т. Маниатисом, нашей лабораторией и др.)
Другой тип семейства - семейство, в котором его представители находятся близко друг от друга в одной области генома, но не образуют гомогенных повторяющихся единиц. Это наиболее распространенный тип организации семейств генов. Классический пример - β-глобиновые гены генома человека (см. рис. 11). В области размером ~ 60 т. п. н. хромосомы 11 располагается 7 копий семейства β-глобиновых генов. Все они несколько отличаются друг от друга и имеют разные названия: ε, γА, γИ, δ и β. Пять копий функциональны и экспрессируются на разных стадиях онтогенеза. Две копии представляют собою псевдогены, или испорченные гены, в которых произошли мутации, не совместимые с функционированием гена.
Очевидно, все гены β-глобинового семейства произошли путем дупликации одного родоначального Р-глобинового гена. В ходе дальнейшей эволюции они приобрели регуляторные последовательности, обеспечивающие их работу на определенной стадии развития: ε - в раннем эмбриогенезе, γ - на более поздних его стадиях, β - в период постнатального развития. Произошло и некоторое изменение их структуры, причем экзоны изменились меньше, чем интроны. В тех генах, которые в ходе эволюции инактивировались, произошло быстрое накопление изменений, образовались псевдогены.
Кроме семейства β-глобиновых генов существует семейство α-глобиновых генов. Они расположены на другой хромосоме и отличаются по структуре, но тем не менее имеют гомологию с β-глобиновыми генами. Очевидно, на еще более ранней стадии эволюции из одного предкового глобинового гена образовались путем дупликации два (α и β), которые затем в результате транслокации (перемещения) генетического материала оказались в двух разных хромосомах. Там они еще несколько раз дупли-цировались, что привело к созданию двух семейств. Вместе они образуют суперсемейство.
Третий тип семейств генов характеризуется тем, что его члены вообще не образуют скоплений, а оказываются разбросанными по геному. Очевидно, в этом случае в промежутке между дупликациями происходили транслокации генов в новые места генома. Сюда, например, относятся представители семейства актиновых генов у дрозофилы.
Семейства генов бывают большие и малые. Так, например, семейство генов для фактора некроза опухолей (ФНО) состоит всего из двух генов - ФНО-α и ФНО-β. Они расположены в геноме рядом с интервалом всего в 1 т. п. н. (см. рис. 11).
В то же время суперсемейство генов иммуноглобулинового типа состоит из нескольких семейств (собственно иммуноглобулиновые, гистосовместимости, Т-клеточных рецепторов и др.) и содержат тысячи копий, занимая огромные участки генома, в миллионы пар нуклеотидов.
В настоящее время литература по семействам очень обширна. Подобное рассмотрение ее выходит за рамки книги. Отмечу лишь, что принцип организации генов в семейства указывает на исключительно важную роль процесса дупликации генов в ходе эволюции. Это один из важнейших путей образования нового генетического материала.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'