2.2.3. Половые хромосомы
2.2.3.1. Первые наблюдения
Нерасхождение половых хромосом и определение пола у дрозофилы. В 1910 г. Морган [448] впервые детально описал X-сцепленное наследование и X-Y-тип хромосомного определения пола у Drosophila melanogaster. В его опытах наблюдалось несколько исключений, не согласующихся с предположением о сцеплении с X-хромосомой. Бриджес объяснил эти исключения, предположив возможность особых аномалий в механизме мейоза. В 1916 г. он показал [311], что в мейозе у дрозофилы действительно может иметь место нерасхождение половых хромосом. Дрозофила имеет четыре пары хромосом: три аутосомы и две половые хромосомы. Так же как и у человека, самки имеют набор XX, а самцы XY, т. е. все самцы по генам, сцепленным с X-хромосомой, являются гемизиготными (полузиготными). Стало быть, каждая нормальная мужская гамета будет нести либо X-, либо Y-хромосому, а все яйцеклетки X-хромосому. В скрещиваниях самок, гомозиготных по X-сцепленному признаку white (белоглазые), с самцами дикого типа (красноглазыми) все потомки-самцы, будучи гемизиготными, должны иметь белые глаза, как и их гомозиготные матери. Все дочери должны быть гетерозиготными и иметь нормальные красные глаза. Как правило, эти предположения оправдывались, однако в очень редких случаях самцы имели нормальные красные глаза, а самки белые. Бриджес показал, что это связано с нерасхождением материнских X-хромосом, что приводит к образованию яйцеклеток либо с двумя X-хромосомами, либо вообще без них (рис. 2.67). После оплодотворения спермиями самца дикого типа возможно образование зигот четырех типов: XXX; XXY; XO; YO. Зиготы YO не обнаружены, очевидно, в силу их нежизнеспособности. Остальные три типа зигот действительно обнаруживаются. Существование таких мух позволяет делать вывод относительно механизма определения пола:
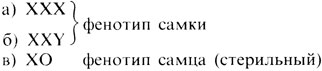
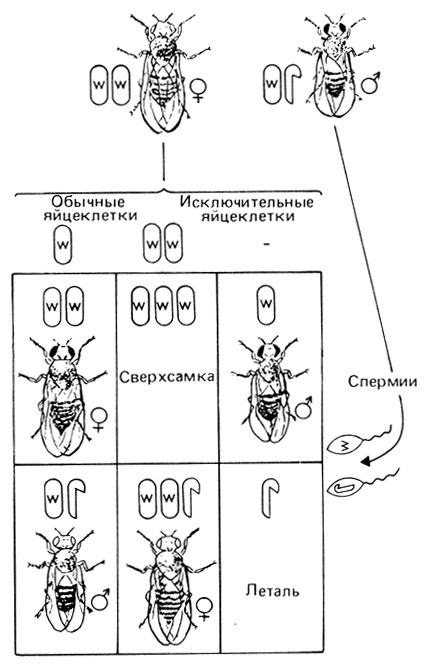
Рис. 2.67. Нерасхождение X-хромосом у Drosophila, скрещивание самок white с самцами дикого типа. (Sinnott-Dunn-Dobzhansky, Principles of Genetics, 1958.)
Следовательно, фенотипический пол у плодовой мушки зависит от числа X-хромосом. Одна C-хромосома определяет пол самца, большее их число пол самки. Y-хромосома также влияет на определение пола, поскольку самцы ХО стерильны.
Генотип XO у мыши. X-сцепленная мутация scurfy (sf) (шерсть животных как бы покрыта пылью) впервые была выявлена как спонтанная. Гемизиготные самцы стерильны, поэтому линию можно поддерживать только путем скрещивания гетерозигот (Xsf/X+) с нормальными самцами (X+/Y). Соотношение мутантных и нормальных самцов в таком скрещивании должно быть 1:1, а все самки в потомстве должны быть нормальными. Однако изредка появляются исключительные самки. Подобно самцам-гемизиготам, они стерильны. Яичники таких самок пересаживали нормальным самкам, которых затем скрещивали с самцами дикого типа. В потомстве от такого скрещивания все самцы были sf, а все самки - нормальные. Фенотипически нормальные самки разделялись, однако, на две группы: одни из них передавали, а другие не передавали "мутацию scurfy своим потомкам. Дальнейший анализ показал, что эти самки различаются генетически: X+/O и X+/Xsf; самки с первым кариотипом не передавали sf в отличие от самок со вторым кариотипом. Этот эксперимент показывает, что особи ХО у мыши являются фертильными самками. Следовательно, у мыши за фенотипический пол ответственна не X-хромосома, а Y-. Впоследствии XO-кариотип у мыши обнаруживали довольно часто. В большинстве случаев это состояние возникает не вследствие мейотического нерасхождения, а в результате потери хромосомы после оплодотворения. При исследовании мутационного процесса утрата хромосомы стала важным инструментом для учета мутагенной активности (разд. 5.2). Позже у мыши был обнаружен и кариотип XXY. В отличие от дрозофилы, у которой особи XXY являются самками, у мыши особи XXY имеют фенотип самцов.
Анеуплоидия по X-хромосоме у человека: XXY; XO; XXX. Анеуплоидия по X-хромосоме была первой из выявленных у человека хромосомных аномалий. Когда Джекобс и Стронг (1959) [395] обследовали 42-летнего мужчину с типичными признаками синдрома Клайнфельтера (рис. 2.68; 2.69) (гинекомастия, маленькие тестикулы, гиалиноз тестикулярной ткани), они обнаружили Х-хроматин в клетках эпителия ротовой полости и "барабанные палочки" в нейтрофильных лейкоцитах. При исследовании хромосом в клетках костного мозга была выявлена добавочная субметацентрическая хромосома "в группе средних по размеру". Авторы предположили, что кариотип больного - XXY. Однако "не может быть исключена возможность... что добавочная хромосома представляет собой аутосому, несущую феминизирующие гены". Оба родителя больного имели нормальный кариотип с 46 хромосомами, следовательно, нерасхождение произошло у одного из них в мейозе. Вскоре после этого кариотип XXY был подтвержден при многих других случаях синдрома Клайнфельтера.
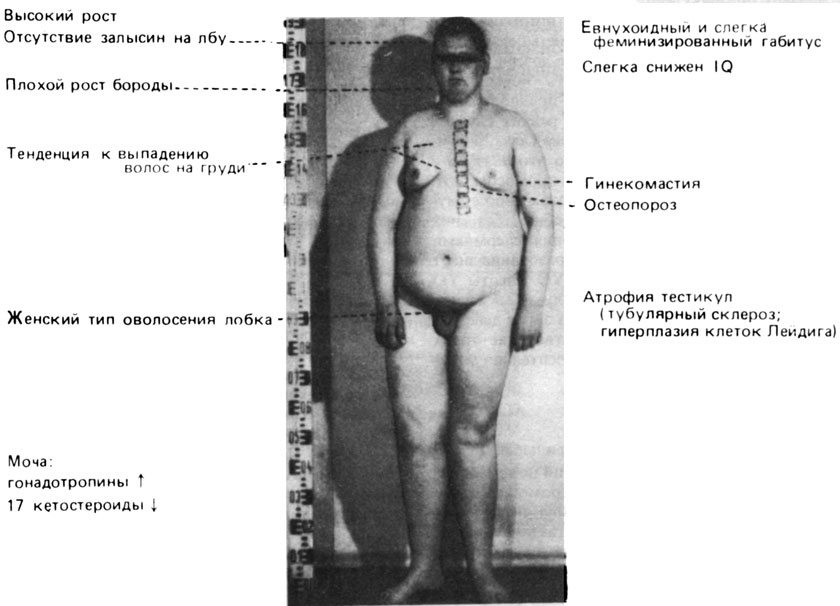
Рис. 2.68. Основные клинические симптомы синдрома Клайнфельтера
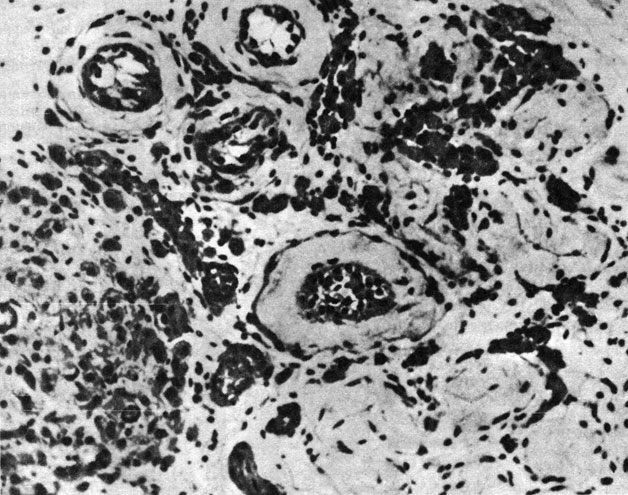
Рис. 2.69. Гиалинизированная тестикулярная ткань при синдроме Клайнфельтера. Нормальные семенные трубочки отсутствуют и замещены гиалинизированной тканью
В то же самое время Форд и сотр. (1959) [352] выявили кариотип ХО. В этом случае 14-летняя девочка имела клинические признаки синдрома Тернера (рис. 2.70) при отсутствии в клетках эпителия слизистой оболочки рта X-хроматина. Модальное число хромосом в клетках костного мозга было 45, обнаружено только 15 "метацентрических хромосом средней длины", как у нормальных мужчин. Это строго соответствовало кариотипу XO. Сравнивая эти результаты с тем, что было известно для дрозофилы (рис. 2.67), авторы пришли к выводу, что в противоположность плодовой мушке тип ХО у человека приводит к развитию "агонадального" индивида с женским фенотипом. Упомянув о кариотипе XXX у дрозофилы, они отметили, что у человека он еще не описан.
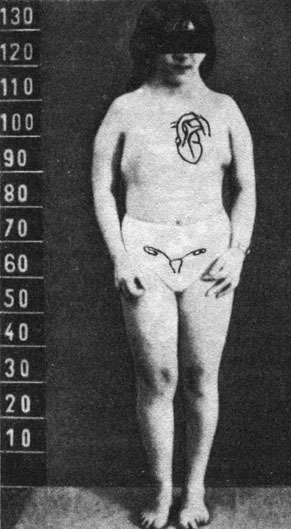
Рис. 2.70. Основные клинические симптомы синдрома Тернера. (Низкая граница роста волос на затылке. Щитовидная грудная клетка. Широко расставленные соски. Укорочение метакарнальных костей IV. Гипоплазия ногтей. Многочесленные пигментные пятна. Низкий рост. Лицо 'сфинкса'. 'Рыбный' рот. Крыловидная складка шеи. Коарктация аорты. Слабое развитие молочных желез. Вальгусное искривление локтя. Рудиментарные яичники. Фиброзный тяж на месте гонад. Первичная аменорея. Дорсальная метакарнальная и метатарсальная лимфедема (при рождении). Моча: гонадотропины ↑, 17 кетостероиды ↓. Эстрогены ↓)
Вскоре, однако, появилось сообщение о 35-летней женщине с плохо развитыми наружными половыми признаками и вторичной аменореей. Ее хромосомный набор состоял из 47 хромосом, причем добавочной была явно X-хромосома: 47, XX, + X. В этом случае были исследованы и костный мозг, и фибробласты. Во многих клетках слизистой оболочки рта и в некоторых нейтрофилах удалось обнаружить два тельца X-хроматина. Таким образом,
- в противоположность дрозофиле фенотипический пол у человека определяется наличием или отсутствием Y-хромосомы, а не числом X-хромосом. В этом отношении человек подобен мыши, но тип ХО у мыши соответствует фертильной самке, в то время как у человека-это женщина с нефункционирующими яичниками;
- число телец X-хроматина на одно меньше, чем число X-хромосом.
Эти два факта стали фундаментом наших знаний и гипотез относительно детерминации пола и генетической активности X-хромосом.
2.2.3.2. X-хромосомные анеуплоидии у человека: современное состояние проблемы
Различие между X-хромосомными и аутосомными анеуплоидиями. Вскоре после первых открытий анеуплоидии по половым хромосомам последовали и другие. Если рассматривать все эти случаи в целом как группу, то обнаруживаются заметные отличия от аутосомных анеуплоидии, о которых шла речь ранее:
- умственное развитие в среднем хотя и ниже нормы, но аномалии развития мозга выражены не столь отчетливо, как при аномалиях аутосом. Многие пробанды имеют нормальное умственное развитие, а некоторые - даже выше среднего (разд. 8.2.2.2);
- фенотипические нарушения в большей степени затрагивают развитие половых органов и гормонозависимый рост. Наблюдаются и другие пороки развития, особенно при синдроме Тернера, но встречаются они реже и по масштабам менее тяжелые.
В конечном счете X-хромосомная анеуплоидия не нарушает эмбриональное развитие в такой степени, как аутосомные анеуплоидии. Объяснение, по-видимому, состоит в том, что в норме женщина имеет две, а мужчина только одну X-хромосому, в связи с чем в эволюции сформировался специальный мощный механизм компенсации различий в дозе генов, сцепленных с X-хромосомой. Этот же механизм оказался фактором, благоприятствующим носителям X-хромосомных анеуплоидии.
Клиническая классификация X-хромосомных анеуплоидии: мозаики. Наиболее важные численные и структурные аномалии X-хромосом представлены в табл. 2.10. В общем, число добавочных X-хромосом увеличивает степень умственной отсталости. Число телец X-хроматина всегда на одно меньше, чем число X-хромосом. В табл. 2.10 описаны также наиболее частые типы мозаицизма, однако обмены с участием X-хромосомы не указаны. В последнем случае приложимы те же правила, что и для аутосомных транслокаций, в частности, в отдельных семьях наблюдается значительная вариабельность фенотипических проявлений.

Таблица 2.10. Численные и структурные X-хромосомные анеуплоидии у человека
Некоторые из X-аутосомных обменов важны для разработки теоретических концепций относительно инактивации X-хромосомы.
Крупные пороки развития обнаруживаются чаще всего при гонадальном дисгенезе, который является наиболее гетерогенным в клиническом и цитогенетическом отношении. Некоторые авторы, исходя из клинических принципов, предполагают возможность подразделения этого синдрома на варианты [88, 477]. Наиболее известная классификация основана на противопоставлении, с одной стороны, простого гонадального агенеза без добавочных симптомов, а с другой - синдрома Тернера с признаками, представленными на рис. 2.70. Однако патогенетические данные слабо или вовсе не коррелируют с таким подразделением на две группы.
Анеуилоидия XO встречается много реже, чем варианты XXY и XXX. Ожидаемые частоты различных типов зигот основаны на закономерностях распределения хромосом по гаметам после нерасхождения в первом или втором делениях мейоза у лиц обоего пола (рис. 2.71). При этом делается допущение, что вероятность оплодотворения любой яйцеклетки спермием X или Y составляет 1/2 и что все спермин (включая и анеуплоидные) участвуют в оплодотворении яйцеклеток. Эти события можно обозначить следующим образом:

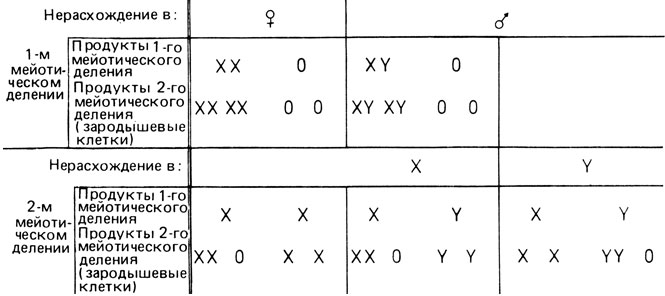
Рис. 2.71. Генотип половых клеток при нерасхождении X- и Y-хромосом в первом и втором мейотическом делении. Подробности см. в тексте
Ожидаемые относительные частоты (вероятности, P) различных типов зигот:
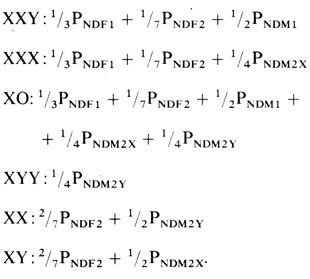
Следовательно, при отсутствии влияния других факторов и при условии равных вероятностей для NDM1 и NDM2X зиготы XXY должны встречаться несколько чаще, чем зиготы XXX. Существуют некоторые доказательства того, что нерасхождение аутосом происходит чаще в первом, чем во втором мейотическом делении (разд. 5.1.2.3).
Теоретически зиготы ХО должны встречаться чаще, чем зиготы других типов. Однако эмпирические данные не соответствуют этому, синдром Тернера встречается много реже, чем зиготы XXX и XXY. Это указывает на наличие сильного отбора против гамет, не содержащих половой хромосомы X, и/или на наличие сильного внутриутробного отбора против зигот XO. Последнее предположение подтверждается тем, что среди спонтанно абортированных зародышей тип ХО встречается относительно часто. Есть и другие доказательства: риск нерасхождения обычно увеличивается с возрастом матери (разд. 5.1.2.2). Для анеуплоидий типа XXX и XXY в отличие от типа XO эта закономерность выявляется вполне четко. В настоящее время допускается, что выжившие зиготы XO являются результатом не мейотического, а митотического нерасхождения или утраты хромосом на ранних стадиях дробления. Об этом же свидетельствуют данные, что в группе больных с синдромом Тернера мозаики встречаются относительно чаще, чем в группах XXX- и XXY-анеусомиков. С другой стороны, зиготы XYY могут возникать только вследствие нерасхождения во втором мейотическом делении у мужчин. Однако они встречаются почти так же часто, как и зиготы XXY. Следовательно, вероятность нерасхождения по Y-хромосоме, PNDM2Y, явно много выше, чем отдельные вероятности разных типов нерасхождения по X-хромосоме. Наблюдались мозаики всех типов. Механизмы возникновения мозаиков обсуждаются в разд. 5.1.6.
Интерсексы. На основании клинических данных различают три типа интерсексов:
- истинный гермафродитизм: присутствуют половые клетки обоих полов;
- мужской псевдогермафродитизм: имеются только тестикулы;
- женский псевдогермафродитизм: имеются только яичники.
К сожалению, эта простая классификация не совпадает с цитогенетическими данными, поскольку в каждой группе встречаются хромосомные варианты, включая и такие случаи, как, например, 46,XX у мужчин. Многие интерсексы являются мозаиками, содержащими клетки с различным набором половых хромосом в разных комбинациях. Например, фенотип мозаиков 45,XX/46,XY может проявиться в виде овариального дисгенеза, гонадального дисгенеза, с мужским псевдогермафродитизмом или в форме "смешанного гонадального дисгенеза", когда одна гонада представлена фиброзным тяжем, а другая - диспластическим тестикулом. Некоторые истинные гермафродиты имеют кариотип 46,XX/46,XY. Мозаицизм типа XX/XY может возникать как следствие любого из девяти различных механизмов, таких, как оплодотворение ооцита двумя различными спермиями, слияние двух оплодотворенных яйцеклеток, митотическая ошибка во время первого дробления или внутриутробный обмен стволовыми кроветворными клетками между разнополыми дизиготными близнецами (разд. 3.8.3).
Первичная функция полоопределяющих факторов состоит в индукции гонад. Гонады в свою очередь детерминируют развитие других половых органов и вторичные половые признаки. Нарушения индукции гонад могут быть вызваны либо аномалиями набора половых хромосом, либо другими факторами, не имеющими прямого отношения к половым хромосомам. В последнем случае интерсексы могут иметь нормальный набор хромосом как XX, так и XY. Сбалансированные структурные перестройки с участием X-хромосомы часто ведут к бесплодию у обоих полов [427].
Y-хромосома как фактор, определяющий мужской пол. Развитие тестикул детерминируется Y-хромосомой. Бюлер [315], обобщив данные многочисленных литературных сообщений о больных с аномалиями половой дифференцировки и структурными аномалиями Y-хромосомы, попытался локализовать различные функции в эухроматическом районе Y-хромосомы человека (рис. 2.72). На этом рисунке отсутствует, однако, информация, полученная на основе использования специфических ДНК-зондов. Различные типы аномалий X-и Y-хромосом весьма информативны относительно механизмов полового развития человека. Однако полезные сведения по этой проблеме, можно получить также и при изучении метаболических аномалий с простым типом наследования, из опытов по получению химер у животных, а также из исследований HY-антигена. Кроме того, для полного понимания процесса половой дифференцировки необходимы знания о генетическом контроле регуляторных функций гормонов. В связи с этим особое значение приобретает концепция рецепторных болезней. Обсуждение этих вопросов будет представлено в разд. 4.7.5, здесь же мы обсудим один специальный вопрос - дозовую компенсацию.
![Рис. 2.72. Слева: функции, аномалии которых отмечены у больных с аберрациями Y-хромосомы [315]. Справа: локализация Y-специфических зондов ДНК (из каталога McKusick 1985)](pic/000089.jpg)
Рис. 2.72. Слева: функции, аномалии которых отмечены у больных с аберрациями Y-хромосомы [315]. Справа: локализация Y-специфических зондов ДНК (из каталога McKusick 1985)
2.2.3.3. Дозовая компенсация X-хромосомы млекопитающих [357]
Природа X-хроматина. Когда Барр и Бертрам (1949) [298] открыли X-хроматин, относительно его природы были высказаны разные гипотезы. По аналогии с дрозофилой вначале предполагалось, что X-хроматин состоит из гетерохроматиновых районов обеих X-хромосом. Это как будто подтверждалось наблюдением, что Х-хроматин состоит из двух частей. Однако Оно и соавт. (1959) [463; 464] показали, что X-хроматин формируется только одной Х-хромосомой. В делящихся диплоидных клетках регенерирующей печени крысы на стадии профазы X-хроматиновое тельце выглядело как одна большая хромосома, сильно конденсированная во всей своей длине, а не как объединение гетерохроматиновых районов двух хромосом. В клетках мужчин такая конденсированная хромосома отсутствовала. Был сделан вывод, что каждое тельце X-хроматина представляет собой единственную гетеропикнотическую X-хромосому. Половой диморфизм наблюдается потому, что единственная X-хромосома у мужчины, как и одна из двух X-хромосом у женщины, остается эухроматической. Этот вывод был подтвержден у других млекопитающих, а Тэйлор (1960) [525] показал с помощью радиоавтографии с меченым 3H-тимидином, что женская гетеропикнотическая X-хромосома в соматических клетках китайского хомячка реплицируется только в конце S-фазы. Результаты Тэйлора были подтверждены исследованиями на многих других млекопитающих. Гетерохроматинизация X-хромосомы происходит на ранних стадиях эмбрионального развития. Дробящаяся зигота млекопитающих не имеет Х-хроматина. Время его первого появления у различных видов варьирует от бластоцисты до стадии ранней первичной полоски, т. е. от 50-клегочной стадии (у свиньи) до стадии в несколько сотен клеток (у человека) и от предимплантационного до постимплантационного периода. В трофобласте человека X-хроматин появляется на 12-й день развития, а в собственно эмбрионе на 16-й день. X-хроматин формируется одновременно во всех клетках эмбриона. Данные об анеуплоидных индивидах, имеющих более двух X-хромосом, свидетельствуют о том, что только одна X-хромосома остается в эухроматическом состоянии, тогда как все остальные гегерохроматинизированы.
Инактивация X-хромосомы как механизм дозовой компенсации: гипотеза Лайон. В 1961 г. Лайон высказала предположение, что в разных клетках одного организма гетеропикнотическая Х-хромосома может быть либо отцовского, либо материнского происхождения и что эта хромосома функционально неактивна. Таким образом, ею была сформулирована одна из наиболее плодотворных гипотез в генетике млекопитающих, связавшая морфологию хромосомы с ее функцией.
Доказательства этой гипотезы опираются на два рода данных. Во-первых, нормальный фенотип самок XO у мыши свидетельствует о том, что для полноценного развития ей необходима только одна активная X-хромосома. Во-вторых, у самок мышей, гетерозиготных по некоторым сцепленным с полом генам, обнаруживается мозаицизм. Так, самки, гетерозиготные по сцепленным с полом мутациям, затрагивающим окраску шерсти, имеют шкурку с пятнами нормальной и мутантной окраски. Этот факт заставляет думать, что мозаичный фенотип в данном случае обязан своим возникновением инактивации одной или другой X-хромосомы еще в эмбриональном развитии. Эта гипотеза предсказывает, что все гены, локализованные в X-хромосоме и находящиеся в гетерозиготном состоянии, будут иметь мозаичное проявление, также будут проявляться и аутосомные гены, транслоцированные на X-хромосому. Когда фенотип не связан с локальным действием гена, возможны различные типы фенотипических распределений. Следовательно, фенотип будет промежуточным между нормальным и гемизиготным, что приведет к неполной пенетрантности у гетерозигот.
В том же году Лайон попыталась на основе своей гипотезы объяснить данные, полученные при изучении заболеваний человека, наследуемых сцепленно с полом: при X-сцеиленном глазном альбинизме у гемизиготного мужчины нет пигмента в эпителиальных клетках сетчатки и глазное дно имеет бледную окраску. У гетерозиготных женщин наблюдается неправильная пигментация сетчатки, с пигментированными и не содержащими пигмента пятнами, так что глазное дно имеет не равномерную окраску, а точечную. На рис. 2.73 изображено такое глазное дно. Лайон предсказала также, что должен существовать мозаицизм и по другим X-сцепленным генам, в частности по вариантам фермента глюкозо-6-фосфат - дегидрогеназы (G6PD).
![Рис. 2.73. Глазное дно 6-летней дочери мужчины с X-сцепленным глазным альбинизмом. Распределение пигментированных пятен у этой гетерозиготы явно неслучайное [8]](pic/000090.jpg)
Рис. 2.73. Глазное дно 6-летней дочери мужчины с X-сцепленным глазным альбинизмом. Распределение пигментированных пятен у этой гетерозиготы явно неслучайное [8]
Доказательство инактивации X-хромосомы по данным генетики G6PD у человека. Окраска шерсти у мыши, контролируемая X-сцепленными генами, или точечная окраска глазного дна при X-сцепленном альбинизме у человека представляют собой фенотипические признаки, отдаленные от первичного действия гена многими этапами дифференцировки, следовательно, трактовка происхождения таких фенотипов неоднозначна. Эти наблюдения могут служить основой для гипотезы об X-инактивации, но их недостаточно для ее доказательства. Критическая проверка такой гипотезы подразумевает использование более простых и менее многозначных ситуаций. Хорошей экспериментальной моделью являются X-сцепленные гены, первичные эффекты которых можно обнаружить уже на уровне белка. Первым X-сцепленным локусом, для которого такой анализ стал возможным, оказался ген G6PD. Действительно, Бейтлер (1962, 1964) [302, 303] независимо от Лайон разработал концепцию инактивации X-хромосомы, исходя из анализа вариантов G6PD у человека. Несмотря на то что женщины имеют две, а мужчины - только одну копию гена G6PD, средний уровень активности этого фермента одинаков у обоих полов, так же как и у индивидов с дополнительными X-хромосомами (XXX, XXY). Следовательно, должен действовать механизм дозовой компенсации. Если женщина гетерозиготна по электрофоретическим вариантам G6PD, то гипотеза случайной инактивации предсказывает, что в одних клетках будет активна X-хромосома с нормальным аллелем, а в других с мутантным. Отсюда следует, что данная отдельная клетка способна экспрессировать только один из двух вариантов фермента. Такой мозаицизм действительно был обнаружен Бейтлером в эритроцитах при помощи остроумного, но непрямого метода [304]. Позже этот феномен был подтвержден многими авторами и различными методами [358, 424]. Один из подходов использует клонирование фибробластов в культуре. В африканской популяции ген G6PD полиморфен и представлен двумя частыми аллелями GdA и GdB. В отдельных клонированных фибробластах негритянки, гетерозиготной по этим аллелям, обнаружены либо GdA, либо GdB варианты (рис. 2.74), но не оба вместе. При обследовании другой женщины, гетерозиготной по недостаточности G6PD, обнаружили аналогичное явление: в одних клонах активность фермента была нормальной, а в других - очень низкой. Еще одно доказательство теории X-инактивации получено при изучении лейомиомы матки у женщины, гетерозиготной по вариантам А и В G6PD. В опухолевой ткани неизменно обнаруживали только один из двух мутантных типов, в то время как в нормальной ткани матки обнаружены оба типа. Эти явления возможны только при наличии трех условий.
![Рис. 2.74. Электрофоретическое разделение G6PD из разрушенных ультразвуком клеток культуры ткани гетерозиготной женщины с генотипом GdAB. Обнаружено два компонента G6PD - GdA и GdB. В клонах, происходящих из единичных клеток, выявляется либо GdA, либо GdB, но не оба одновременно [93]](pic/000091.jpg)
Рис. 2.74. Электрофоретическое разделение G6PD из разрушенных ультразвуком клеток культуры ткани гетерозиготной женщины с генотипом GdAB. Обнаружено два компонента G6PD - GdA и GdB. В клонах, происходящих из единичных клеток, выявляется либо GdA, либо GdB, но не оба одновременно [93]
- Активен только один аллель.
- Опухоль начиналась с единственной клетки, т. е. она представляет собой клон единичной клетки.
- Индивидуальные X-хромосомы остаются либо активными, либо неактивными в течение всего периода роста опухоли.
Эти данные подтверждают не только гипотезу Лайон, но и концепцию моноклонального происхождения опухолей. Эксперименты с отдельными клетками были проведены и в отношении X-сцепленных аномалий ферментов, например в отношении недостаточности по гипоксантин-гуанин - фосфорибозилтрансферазе (HPRT). И в этом случае феномен случайной инактивации X-хромосомы был подтвержден. Генетический дефект по HPRT использовался в качестве модели для изучения механизмов действия гена у человека во многих аспектах, что подробнее будет обсуждаться в разд. 4.2.6.
Особенно информативным оказалось обследование женщин, гетерозиготных одновременно по G6PD и по другому X-сцепленному ферменту, фосфоглицераткиназе (PGK) [303]. Было изучено 56 отдельных клеточных клонов, в 33 из них обнаружены фенотипы GdA и PGK-1, в то время как в 23 остальных - GdB и PGK-2. Если бы инактивация этих двух локусов происходила независимо друг от друга, то следовало бы ожидать появления клонов с фенотипом GdA, PGK-2 или GdB, PGK-1.
Другие примеры инактивации X-хромосомы у человека. Инактивация X-хромосомы у человека была продемонстрирована для многих X-сцепленных признаков и с помощью разных методов. Особенно интересна демонстрация мозаицизма сетчатки при цветовой слепоте красно-зеленого типа [309]. Сетчатку женщины, гетерозиготной по цветовой слепоте, освещали узким пучком красного или зеленого света. При этом, как и ожидалось, были обнаружены пятна дефектного цветовосприятия, поскольку сетчатка имела мозаичное строение из нормальных и дефектных клонов.
Ангидротическая эктодермальная дисплазия является редким X-сцепленным заболеванием. У пораженных мужчин нет зубов, потовых желез, имеется гипотрихоз, тогда как у гетерозиготных женщин выявляются участки кожи как нормальные, так и лишенные потовых желез [470].
При хроническом грануломатозе с нарушением функции лейкоцитов (30640) [714] бактерицидная активность гранулоцитов резко снижена: они нормально поглощают стафилококки, но не переваривают их. У гетерозиготных женщин имеются две популяции лейкоцитов - нормальная и аномальная [480, 542]. При других X-сцепленных заболеваниях также обнаружены явления, предсказываемые гипотезой Лайон [133].
Клетки, в которых вторая X-хромосома не инактивируется [424, 426]. Тельце X-хроматина становится видимым примерно на 16-й день, на стадии бластоцисты, и вряд ли функциональная инактивация X-хромосомы происходит раньше. Если одна из X-хромосом с самого начала была неактивной, то различия в фенотипах нормального мужчины (XY) и больного с синдромом Клайнфельтера (XXY), а также нормальной женщины (XX) и больной с синдромом Тернера (XO) нуждались бы в ином объяснении, чем предполагаемая Лайон возможность действия генов X-хромосом до инактивации. Имеются доказательства того, что в ооцитах, так же как и в сперматозоидах, X-хромосома не инактивируется. У мыши фермент LDH контролируется аутосомным геном, а G6PD, так же, как и у человека, - X-сцепленным геном. В оплодотворенных ооцитах ХО была обнаружена сниженная наполовину (по сравнению с XX-ооцитами) активность G6PD, а активность LDH была одинаковой [345]. Отсутствие эффекта дозовой компенсации можно объяснить только исходя из предположения, что в XX-ооцитах обе X-хромосомы являются активными.
Одна из систем групп крови человека, система Xg, наследуется сцепленно с полом. Согласно предсказаниям гипотезы Лайон, следовало бы ожидать, что у гетерозиготных женщин имеются две популяции эритроцитов, в каждой из которых клетки несут антиген, детерминированный той X-хромосомой, которая оказалась активной в клетке-предшественнице данной популяции. Однако вскоре стало ясно, что это предсказание неверно, поскольку не было обнаружено двух разных популяций эритроцитов. Дополнительно допускалась еще возможность того, что антиген Xg образуется не самими эритроцитами, а привносится их окружением, например сывороткой. И это предположение было опровергнуто наблюдениями на химерных близнецах (разд. 3.8.3). Речь идет о женщинах, которые в дополнение к своим собственным эритроцитам получили стволовые клетки крови от их дизиготных близнецов во время эмбрионального развития. Одни эритроциты имели антигены O и Xga+, другие были AB и Xga-. Если бы Xg транспортировался из сыворотки, все клетки имели бы тип Xg - генетически "собственный" тип пробанда. Эта загадка была раскрыта, когда установили, что локус Xg расположен вблизи конца короткого плеча X-хромосомы и что по крайней мере еще один локус, тесно сцепленный с Xg фермента стероидсульфатазы, также не вовлекается в инактивацию. Иначе говоря, дистальная часть короткого плеча X-хромосомы человека не инактивируется [486, 487]. Активный район охватывает, по-видимому, сегменты Xp 22.13 и Xp 22.3 [502]. В отличие от всех остальных районов инактивированной X-хромосомы он реплицируется в ранней S-фазе.
Что происходит раньше, инактивация X-хромосомы или образование X-хроматина? На первый взгляд представляется очевидным, что сначала X-хромосома конденсируется с образованием X-хроматина, а затем прекращает функционировать. На самом деле ход событий скорее всего противоположный: сначала происходит инактивация, затем формируется X-хроматин. Такой вывод следует из того факта, что X-хроматин никогда не обнаруживается одновременно во всех клетках. Например, в клонированных культурах фибробластов примерно 30% клеток не имеют X-хроматина. Количество клеток с X-хроматином зависит, по-видимому, от клеточного цикла. Измерения активности G6PD в таких культурах показали, что функциональная инактивация была практически полной и отсутствовало какое-либо соответствие между активностью фермента и числом телец X-хроматина в фибробластах от индивидов с различным числом X-хромосом. Точный механизм инактивации является еще предметом исследований [357].
Существуют ли генетические различия в паттернах X-инактивации? Каттанах (1975) [321] описал у мыши X-сцепленный ген, который контролирует инактивацию X-хромосомы (X chromosome controlling element, Xce). Существует мутантный аллель этого гена "выраженная мозаичность" (high variegation, Ohv), который заставляет X-хромосому оставаться в активном состоянии. Исходя из этого наблюдения, можно предполагать, что и у человека инактивация X-хромосомы может находиться под генетическим контролем. Неслучайная инактивация должна иногда приводить к появлению гетерозигот с клиническими признаками X-сцепленных рецессивных болезней. Если инактивация происходит достаточно рано во время эмбрионального развития - в то время, когда количество клеток данной ткани еще довольно невелико, - то и в этом случае должны иногда появляться пораженные гетерозиготы. Они являются крайними вариантами, которые образуют "хвост" биномиального распределения всех паттернов инактивации. Однако гипотеза случайной инактивации не предсказывает накопления таких случаев среди сибсов. Тем не менее накопление наблюдалось в случае мышечной дистрофии Дюшенна [451] и в одной семье со сфинголипидозом (болезнью Фабри) [488]. В этой семье девять гетерозиготных дочерей больного мужчины можно было разделить на два класса: в одной группе у четырех дочерей активность α-галактозидазы А достигала 50%, в то время как в другой группе активность составляла 20% (активность определяли в лейкоцитах). Авторы обсуждают гипотезу, согласно которой имеется ген, детерминирующий предпочтительную инактивацию X-хромосомы с нормальным аллелем. Случаи гетерозиготного проявления мышечной дистрофии Дюшенна можно, вероятно, объяснить таким же образом. Точное определение генной активности у гетерозигот по X-сцепленным болезням способствует накоплению и обобщению подобных сведений.
X-инактивация и аномальные X-хромосомы [529]. Когда были описаны первые аномальные X-хромосомы у человека (изохромосомы по длинному плечу, кольцевые хромосомы или делеции части длинного плеча), правила инактивации казались простыми: всегда инактивируется аномальная X-хромосома, в клетке остается одна нормальная активная X-хромосома. Чтобы объяснить столь специфический характер инактивации, были выдвинуты две гипотезы. В соответствии с селекционной гипотезой нормальная и аномальная X-хромосомы инактивируются случайно, так же как и в случае, когда обе нормальные. Однако клетки с инактивированной нормальной X-хромосомой оказываются генетически несбалансированными в большей степени и поэтому должны иметь более низкую скорость деления, чем те клетки, в которых инактивирована аномальная X-хромосома и которые являются по сути нормальными. Вторая гипотеза предполагает, что инактивация - облигатное свойство аномальных X-хромосом [156].
По мере накопления данных об аутосомных или X/X транслокациях становилось очевидным, что ни одна из этих гипотез не может быть полностью верной. Существует три группы таких транслокаций: реципрокные сбалансированные, практически все они X-аутосомного типа, причем общее число хромосом равно 46; несбалансированные X-аутосомные и X/X транслокации, также при наличии 46 хромосом; несбалансированные X-аутосомные транслокации с общим числом хромосом 45. Мы обсудим данные по первой группе, поскольку для остальных эти данные в основном подтверждают выводы, сделанные для транслокаций первой группы.
В большинстве случаев таких транслокаций нормальная X-хромосома инактивируется. Фенотип проявляется в виде гонадального дисгенеза, иногда в сочетании со слабовыраженными признаками синдрома Тернера. Были описаны семьи, в которых у одного из носителей перестройки инактивирована нормальная, а у другого аномальная X-хромосома. Например, в одной семье у матери обнаружена сбалансированная транслокация X/21 (рис. 2.75). Одна транслокационная хромосома состояла из длинных плеч, другая из коротких плеч X-хромосомы и хромосомы 21 с точкой разрыва вблизи от центромеры (но неясно, с какой стороны). У матери, судя по поздней репликации, инактивированной оказалась нормальная X-хромосома. В клетках этой женщины присутствовало одно тельце X-хроматина. В то же время у ее дочери имелась не маленькая, а большая транслокационная хромосома и две нормальные X-хромосомы. Одна из них была инактивирована, но в отличие от матери транслокационная хромосома тоже была инактивирована. Следовательно, дозовая компенсация достигалась у матери и дочери сходным образом. Однако у дочери инактивация распространялась за пределы X-хромосомы на транслоцированное длинное плечо хромосомы 21, что привело к появлению дополнительных клинических признаков, сходных с теми, которые иногда описываются при моносомии 21.
![Рис. 2.75. X-дозовая компенсация у матери и дочери с разными наборами X-хромосом. У матери две транслокационные хромосомы X-21 и одна нормальная (она и инактивируется); у дочери две нормальные X-хромосомы, одна из которых инактивирована, и одна транслокационная хромосома, которая тоже инактивирована [501]](pic/000092.jpg)
Рис. 2.75. X-дозовая компенсация у матери и дочери с разными наборами X-хромосом. У матери две транслокационные хромосомы X-21 и одна нормальная (она и инактивируется); у дочери две нормальные X-хромосомы, одна из которых инактивирована, и одна транслокационная хромосома, которая тоже инактивирована [501]
Этот случай показывает, что гипотеза, согласно которой инактивация определяется структурой аномальной X-хромосомы, в общем случае неверна. В этой и других семьях, в которых был обнаружен только один инактивационный паттерн, клетки оставались генетически относительно сбалансированными. Примечательно то, что у большинства носителей сбалансированных транслокаций с участием X-хромосомы имеется аномальный фенотип, тогда как носители сбалансированных транслокаций между аутосомами обычно имеют нормальный фенотип. Возможны два объяснения. Либо непрерывность определенного района длинного плеча X-хромосомы необходима для завершения дифференцировки по женскому типу, и в этом случае дефектный фенотип должен быть связан с явлением, называемым в экспериментальной генетике эффектом положения, либо инактивация одной и той же X-хромосомы во всех клетках вызывает эффект, зависящий, вероятно, от функциональной гемизиготности рецессивного гена. В разных случаях, вероятно, действует тот или иной механизм.
У некоторых больных с несбалансированными транслокациями X-хромосом обнаруживается двойное тельце X-хроматина, чего никогда не бывает у нормальных индивидов. Это еще одно наблюдение, относящееся к инактивации X-хромосомы. Описано мною изохромосом по длинному плечу i(Xq) и в то же время лишь единичные случаи i(Xp), хотя оба типа должны встречаться с одинаковой частотой, так как возникают вследствие аномального деления центромеры [519]. С другой стороны, известны Xq-делеции. Эти факты легли в основу гипотезы, согласно которой предполагается существование инактивационного центра в проксимальной части длинного плеча X-хромосомы. Если этот центр присутствует в аномальной хромосоме, то она инактивируется. Если два центра, как при некоторых несбалансированных транслокациях X-хромосомы, то могут образоваться два тельца X-хроматина. Если центра нет, как в большинстве i (Xp)-изохромосом. инактивация не может произойти и зигота, будучи функционально трисомной по короткому плечу X-хромосомы, становится крайне несбалансированной и неспособной к развитию. Недавно получены данные о том, что инактивационный центр может быть расположен на границе между проксимальным Q-темным и Q-светлым сегментами (~ql3) [530]. По некоторым наблюдениям инактивационный импульс, создаваемый этим центром, может распространяться по длине X-хромосомы в направлении короткого, но не длинного плеча. На рис. 2.76 показаны различные типы аномальных X-хромосом, их инактивационный паттерн и фенотипы. Относительно X-аутосомных транслокаций имеется намного больше сообщений, касающихся мышей, чем человека. Некоторые данные по этому вопросу также подкрепляют гипотезу о возможном существовании инактивационного центра. Имеются, в частности, указания на то, что можно даже усилить или ослабить с помощью отбора степень инактивации аутосомного сегмента, транспонированного на X-хромосому. Относительно молекулярных механизмов инактивации X-хромосомы есть много гипотез, однако пока какие-либо определенные выводы преждевременны [357].
![Рис. 2.76. Моносомия, дисомия и трисомия по разным частям X-хромосомы человека, их инактивационные паттерны и фенотипические эффекты [529]. Длинные плечи изображены черными, короткие плечи белыми](pic/000093.jpg)
Рис. 2.76. Моносомия, дисомия и трисомия по разным частям X-хромосомы человека, их инактивационные паттерны и фенотипические эффекты [529]. Длинные плечи изображены черными, короткие плечи белыми
Происходит ли инактивация X-хромосомы в сперматогенезе? Лифшиц и Линдслей (1972) [420] выдвинули гипотезу, согласно которой X-хромосома инактивируется не только у индивидов, имеющих их несколько, но также и в первичных сперматоцитах во время нормального сперматогенеза. Вполне возможно, что это необходимо для нормального созревания сперматозоидов. Больные мужского пола с синдромом Дауна бесплодны из-за остановки сперматогенеза. При исследовании стадии пахитены в сперматогенезе у больных с синдромом Дауна обнаружено, что дополнительная хромосома 21 конъюгирует с X-Y комплексом [399]. Для объяснения некоторых фактов, полученных на мышах, также необходимо допустить, что мейотическая конъюгация аутосом или их плеч со свободной частью X-хромосомы должна быть обычным явлением, и это может приводить к угнетению инактивации X-хромосомы и созревания сперматозоидов.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'