4.1.1. Селективный анализ гаплоидных рекомбинантов
Предварительное установление сцепления. Селективный анализ гаплоидных рекомбинантов - основной и сравнительно простой метод картирования хромосомы у стрептомицетов, с помощью которого можно получить информацию о последовательности маркеров на генетической карте и кольцевой структуре генома. На первом этапе картирования можно скрещивать два двойных ауксотрофных мутанта, а также проводить 5-7-факторные скрещивания, однако результаты последних сложнее анализировать. После роста смешанной культуры на полноценной среде споры высевают на селективные среды для выделения гаплоидных рекомбинантов. При 4-факторном скрещивании ab + + Х + + cd высев спор смешанной культуры проводится на четыре разные селективные среды, отбирающие рекомбинанты по разным парам селектируемых маркеров (по одному от каждого родителя). Из 14 возможных генотипов рекомбинантов на селективных средах вырастает 9 генотипов, в том числе две комплементарные пары генотипов (табл. 4.2).

Таблица 4.2. Селективный анализ гаплоидных рекомбинантов
(Примечание. (+) и (-)соответственно наличие и отсутствие роста рекомбинантов.)
Как видно из табл. 4.2, некоторые генотипы можно выделить на двух или даже на всех четырех селективных средах и их число должно быть приблизительно одинаковым, допускающим случайный разброс. Если члены двух пар комплементарных генотипов представлены приблизительно одинаковыми частотами, то можно предположить почти равный вклад генетического материала обоих родителей в образование зиготы и на этом основании принять частоту любого рекомбинантного генотипа за частоту его комплементарной пары. Тогда общий анализ всех изолированных генотипов дает возможность установить относительное сцепление между всеми парами маркеров на кольцевой генетической карте. Далее можно подсчитать среднее число генотипов, выделенных на разных средах, а затем определить относительные частоты рекомбинаций (перекрестов) в каждом из четырех интервалов карты. Нужно иметь в виду, что полученные частоты рекомбинации относительны из-за отсутствия частоты пары потомков, имеющих родительские комбинации маркеров. Наименьшие частоты рекомбинации между парой маркеров могут указывать на их сцепление на карте.
Следующий этап генетического анализа состоит в установлении характера сегрегации на каждой среде неселектируемых маркеров относительно друг друга. Независимая сегрегация неселективных маркеров указывает на отсутствие их сцепления, а зависимая - на сцепление. Отношение между маркерами легко можно установить с помощью таблицы 2x2, составленной для каждой среды (рис. 4.1). Из рис. 4.1. видно, что сегрегация аллелей а+/а независима от сегрегации аллелей в то время как сегрегация с+/с зависима от сегрегации d+/d, что указывает на сцепление между локусами с и d на кольцевой карте генома.
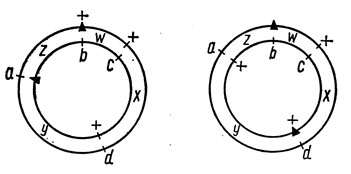
Рис. 4.1. Определение сцепления между локусами с помощью табл. 2 X 2:1 - неселектируемые аллели расположены в той же дуге между селектируемыми маркерами (треугольники) и сегрегируют зависимо друг от друга; 2 - неселектируемые аллели расположены в разных дугах по отношению к селектируемым и сегрегируют независимо; а, b, с, d - маркеры; х, у, w, z - интервалы карты
Картирование новых маркеров. После установления последовательности ряда маркеров на кольцевой карте генома локализация новых маркеров осуществляется с помощью простого селективного метода. В основе его лежат два главных критерия: градиент частот аллелей родителей в рекомбинантном потомстве и минимум множественных кроссинговеров, необходимых для образования наблюдаемых типов рекомбинантов. Рассмотрим применение указанных критериев картирования на примере скрещивания штаммов 97 ade-1 leu-2 X 204-76 сса-5 inl-2 arg-1 Streptomyces olivaceus VKX [3]. Предположим, что нам нужно картировать маркер arg-1. При использовании селектируемых маркеров leu+ и сса+ было выделено 100 гаплоидных рекомбинантов, распределившихся на 6 разных генотипов (табл. 4.3). Вклад аллелей родителей в рекомбинантное потомство представлен на рис. 4.2. По градиентам частот аллелей можно построить две модели расположения маркера arg-1: между локусами leu-2 - inl-2 и leu-2 - ade-1. Выбор между моделями можно сделать на основании минимума множественных кроссинговеров. Как видно из табл. 4.3, модель 1 предусматривает отсутствие множественных кроссинговеров для образования наблюдаемых генотипов рекомбинантов, в то время как согласно модели 2 требуется 25 четверных кроссинговеров для возникновения прототрофных рекомбинантов. Предпочтительна первая модель.
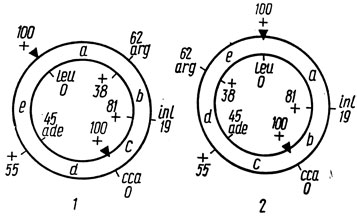
Рис. 4.2. Частота аллелей гаплоидных рекомбинантов, выделенных на селективной среде (треугольники - селектируемые аллели), и модели (1, 2) расположения локуса arg1. Данные табл. 4.3
![Таблица 4.3. Анализ гаплоидных рекомбинантов в скрещивании штаммов 97 ade-1 leu-2 X 204 сса-5 inl-2 arg-1 [3]](pic/000067.jpg)
Таблица 4.3. Анализ гаплоидных рекомбинантов в скрещивании штаммов 97 ade-1 leu-2 X 204 сса-5 inl-2 arg-1 [3]
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'