4.1.2. Неселективный анализ гаплоидных рекомбинантов
Ультрафертильные скрещивания определенных штаммов стрептомицетов впервые у бактерий открывают возможность проведения анализа рекомбинантного потомства в неселективных условиях. У S. coelicolor A3 (2) неселективный анализ гаплоидных рекомбинантов можно проводить в скрещиваниях штаммов NF X UF, в которых выход рекомбинантов достигает частоты 100 %. У S.olivaceus VKX также были выделены ультрафертильные штаммы pSOl-ant, потерявшие плазмиду pSOl и способность синтезировать антибиотик. В скрещиваниях данных штаммов со штаммами pSOl ant, наблюдается выход рекомбинантов с частотой 50-100 %, что позволяет выделять рекомбинанты непосредственно из полноценной среды. В табл. 4.4 приведен неселективный анализ гаплоидных рекомбинантов у S. olivaceus VKX. Картирование маркеров проводилось на основании градиента частот аллелей родителей в рекомбинантном потомстве и минимума множественных кроссинговеров. Как видно из рис. 4.3, в приведенных скрещиваниях в отличие от селективного анализа наблюдается однонаправленный (по часовой стрелке) градиент частот аллелей родителей, максимум которого связан с локусом ant - предполагаемым началом переноса хромосомы.
![Таблица 4.4. Неселективный анализ гаплоидных рекомбинантов в скрещивании штаммов 196 ade-1 his-1 ura-1 (pSO1+) и 24-141 leu-1 rib-1 met-1 ant (pSO1-) [3]](pic/000068.jpg)
Таблица 4.4. Неселективный анализ гаплоидных рекомбинантов в скрещивании штаммов 196 ade-1 his-1 ura-1 (pSO1+) и 24-141 leu-1 rib-1 met-1 ant (pSO1-) [3]
(Примечание. Все колонии образуют зоны подавления роста штамма 585 UF S. coelicolor, т. е. относятся к типу ant+ (pSOl+).)
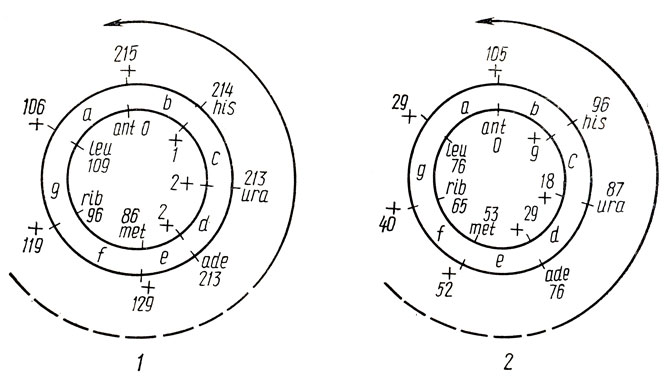
Рис. 4.3. Градиент частот аллелей гаплоидных рекомбинантов и направления переноса хромосомы (стрелка) в скрещиваниях штаммов 196 X 24-141 (pSO1- ): а - g - интервалы карты; 1 - опыт 1; 2 - опыт 2
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'