7.2.1. Хромосомная эволюция и видообразование
Число хромосом у людей и близкородственных им приматов [1912]. Главное внимание мы уделим таким видам, как шимпанзе (Pan troglodytes), карликовый шимпанзе (Pan paniscus), горилла (Gorilla gorilla), орангутан (Pongо pygmaeus). Число хромосом у всех четырех видов равно, как это было давно установлено, 48; основное различие между человеком и обоими видами шимпанзе состоит в наличии у последних дополнительной четвертой пары акроцентрических хромосом группы D (рис. 7.3). У других двух видов - гориллы и орангутана - обнаружено еще больше акроцентрических хромосом. Выявлено убедительное сходство хромосом Homo и Pan, что подтверждает данные морфологических и биохимических исследований (разд. 7.2.3), согласно которым шимпанзе - наш ближайший из ныне живущих филогенетический родственник.
![Таблица 7.2. Эволюция человека [1968]](pic/000007.jpg)
Таблица 7.2. Эволюция человека [1968]
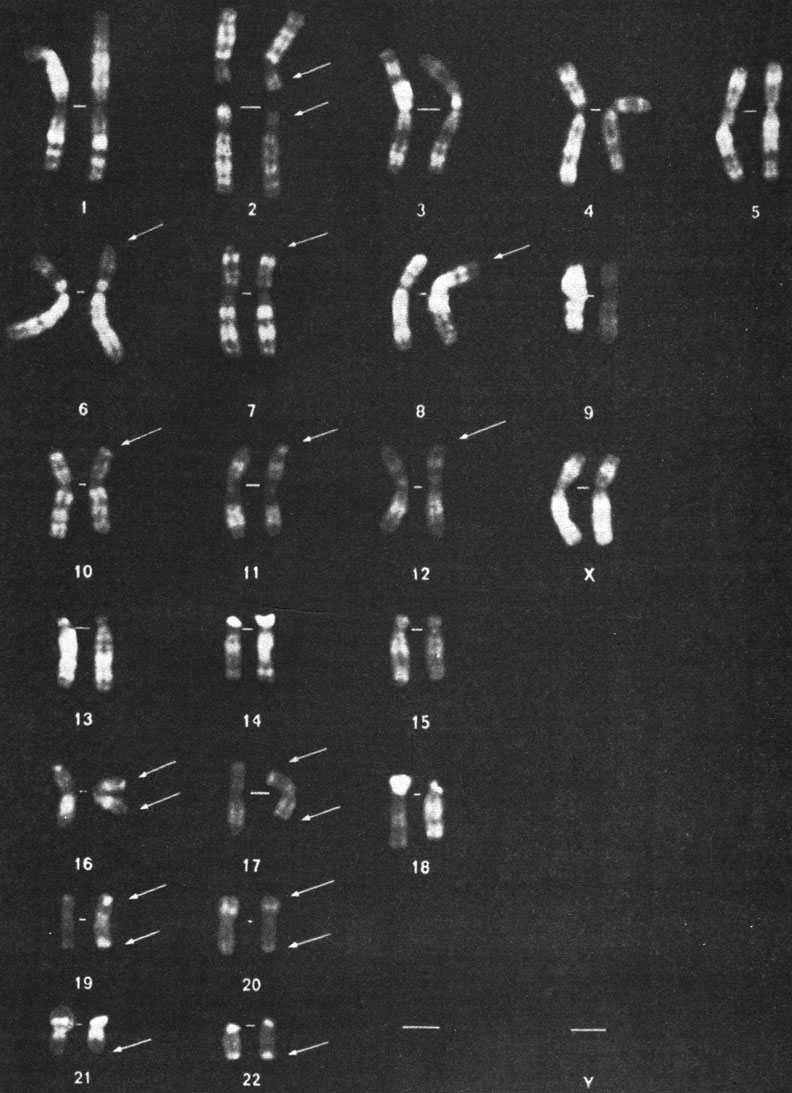
Рис. 7.3. Кариотип шимпанзе (Pan troglodytes), Q-окраска. Хромосома, соответствующая 2-й хромосоме человека, очевидно, отсутствует. Вместо нее имеются две дополнительные пары акроцентрических хромосом. Стрелки указывают на терминальные Q-сегменты, отсутствующие у человека (и орангутана). (Courtesy of Dr. В. Dutrillaux, Paris.) Приведенные числа соответствуют нумерации хромосом человека
Сравнение структуры хромосом с помощью методов дифференциального окрашивания. Сравнение кариотипов двух видов должно помогать в реконструкции числа и типа хромосомных перестроек, произошедших после расхождения этих видов в ходе эволюции. Такая реконструкция стала возможной после разработки в 1970 г. методов дифференциального окрашивания (разд. 2.1.2.2). Вскоре было установлено, что механизмы, ответственные за большинство видовых различий между человеком и человекообразными обезьянами, связаны с перицентрическими инверсиями [1929]. Различие в числе хромосом можно объяснить объединением двух различных акроцентрических хромосом, по длине примерно равных D-хромосомам. Такое событие, вероятно, привело к образованию одной длинной субметацентрической хромосомы - 2-й хромосомы человека. Хорошо известно, что объединение разных хромосом имеет место и в современной человеческой популяции: как правило, это происходит при центрическом слиянии, которое сопряжено с потерей коротких плеч хромосомы. Вполне возможно, что именно этот феномен является причиной обнаруженных видовых различий. Детальный анализ результатов дифференциального окрашивания показал, что материал коротких плеч имеется. Хромосома номер 2 человека возникла в результате теломерного слияния [1912]. Подобного рода события приводят к формированию хромосом, имеющих две центромеры (дицентриков), что в свою очередь сопровождается нарушениями митоза, аналогичными тем, которые индуцируются радиацией или являются результатами обменов, следующих за разрывами хромосом (разд. 5.2.1.1). Митоз может идти нормально только в том случае, если функционирует лишь одна из центромер дицентрика. Такой феномен действительно наблюдался в случае хромосомных аберраций, возникающих в настоящее время.
Детальный анализ всех визуально обнаружимых перестроек, по которым виды антропоидов отличаются друг от друга и от человека, был проведен Дютрилло (1975) [1912].
Пример. На рис. 7.4 представлены фотографии гомологов 2-й хромосомы человека. Хромосомы Ропдо и Gorilla отличаются инверсией в 2q, a Gorilla и Pan другой инверсией, локализованной в 2р. Теломерное слияние должно было произойти после разделения предков Pan и Homo. На некоторых препаратах 2-й хромосомы человека видна вторичная перетяжка в точке слияния (2qh). Изредка (очень редко) наблюдается эндоредупликация сегментов, соответствующих бывшей 2q-хромосоме, что свидетельствует об известной независимости партнеров, слившихся друг с другом. На основе этих результатов можно реконструировать ход эволюции 2-й хромосомы.
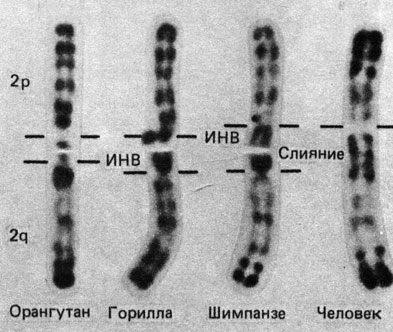
Рис. 7.4. Эволюционные преобразования - несколько инверсий и одно теломерное слияние - в ходе филогенеза 2-й хромосомы человека. G-окраска. Ропдо и Gorilla отличаются инверсией (ИНВ) в 2q; Gorilla и Pan отличаются инверсией в 2р. Homo отличается от всех трех видов теломерным слиянием двух хромосом. (Courtesy of Dr. В. Dutrillaux, Paris.)
Сравнение кариотипов пяти видов. Межвидовые различия использованы для реконструкции эволюции всех хромосом. Кроме одного теломерного слияния и перицентрических инверсий было выявлено несколько парацентрических инверсий. В табл. 7.3 приведены числа хромосомных перестроек разного типа, по которым отличаются сравниваемые виды. Как и ожидалось, наибольшее сходство обнаруживают два вида шимпанзе. Ближайшим родственником Homo является шимпанзе, а самым далеким - Pongo. К такому же выводу можно прийти на основании морфологических данных.
![Таблица 7.3. Сопоставление количества структурных различий* между кариотипами человека и человекообразных обезьян** (числа в скобках - вероятные, но не подтвержденные события) [81]](pic/000010.jpg)
Таблица 7.3. Сопоставление количества структурных различий* между кариотипами человека и человекообразных обезьян** (числа в скобках - вероятные, но не подтвержденные события) [81]
* (Без учета добавочных полос и гетерохроматинового материала.)
** (По данным Дютрилло (1975) [1912].)
Противоречия в полученных данных и их возможное объяснение с неортодоксальных позиций. Для каждой отдельной хромосомы можно установить эволюционные взаимосвязи между различными перестройками, имеющимися у разных видов, и выяснить их филогению. Однако, когда эти хромосомные филогении были совмещены друг с другом с целью реконструкции общей филогении, возникло неожиданное затруднение. Оказалось, что Pan troglodytes и Gorilla имеют три общие инверсии (5, 12 и 17), что свидетельствует о наличии у них общего предка, не являвшегося предком Homo; с другой стороны, Homo и Pan имеют две общие инверсии (2, 7), не найденные у Gorilla. Последний результат свидетельствует о существовании общего предка этих двух видов, не являющегося предком Gorilla. Как разрешить эту дилемму? Дютрилло (1975) [1912] предложил три возможных объяснения данного противоречия (рис. 7.5):
![Рис. 7.5. Три гипотезы, объясняющие противоречия, возникающие при построении филогенетического древа Homo sapiens, Pan troglodytes и Gorilla. А. Гетерозиготность популяции по двум инверсиям. Б. Параллельная фиксация трех инверсий в филогенетических линиях Gorilla и Pan. В. Отделение линий Gorilla и Pan от линии человека; повторяющаяся гибридизация между популяциями предков Gorilla и Pan, которая делает возможной фиксацию в гибридной популяции трех хромосомных перестроек. Впоследствии эти две линии окончательно разделяются. Данная гипотеза считается наиболее вероятной [1912]](pic/000011.jpg)
Рис. 7.5. Три гипотезы, объясняющие противоречия, возникающие при построении филогенетического древа Homo sapiens, Pan troglodytes и Gorilla. А. Гетерозиготность популяции по двум инверсиям. Б. Параллельная фиксация трех инверсий в филогенетических линиях Gorilla и Pan. В. Отделение линий Gorilla и Pan от линии человека; повторяющаяся гибридизация между популяциями предков Gorilla и Pan, которая делает возможной фиксацию в гибридной популяции трех хромосомных перестроек. Впоследствии эти две линии окончательно разделяются. Данная гипотеза считается наиболее вероятной [1912]
1. Общая предковая популяция, возможно, имела хромосомный полиморфизм по этим перестройкам. Однако этот полиморфизм должен был существовать в течение длительного времени, вероятно, на протяжении миллионов лет. Кроме того, мы знаем, что перицентрические инверсии, захватывающие такой большой район хромосомы, как в этом случае, могут приводить к нарушениям мейоза и, следовательно, влиять на репродукцию. Отбор против гетерозигот, который здесь действует, не приводит к стабильному генетическому равновесию (разд. 6.2.1.4). С другой стороны, в Финляндии зафиксирована высокая концентрация перицентрических инверсий, частоты которых почти удовлетворяют критерию полиморфизма (разд. 2.2.2).
2. Второе объяснение сводится к предположению, что в филогенетических линиях Gorilla и Pan troglodytes произошла независимая фиксация трех идентичных перестроек. Однако это событие маловероятно. Поэтому наиболее правдоподобным Дютрилло считает третье из возможных объяснений.
3. Между популяциями предков Gorilla и Pan, после их отделения от предков человека, время от времени происходила гибридизация. Возможно, что вначале произошло разделение предков Gorilla и общих предков шимпанзе и человека, которое сохранялось, вероятно, благодаря экологической изоляции. Много позже разделились линии шимпанзе и человека; предки Pan вселились в ту же экологическую нишу, в которой уже обитала протогорилла (вероятно, эта ниша приурочена к влажным тропическим лесам). Здесь могла происходить повторная гибридизация, продолжавшаяся до тех пор, пока эти два вида не выработали, наконец, репродуктивные барьеры, чем и завершился процесс видообразования.
Другое труднообъяснимое видовое различие - это наличие или отсутствие определенных хромосомных сегментов.
Присутствие или отсутствие определенных сегментов. Помимо описанных выше перестроек у некоторых из этих приматов регулярно выявляются определенные хромосомные сегменты, отсутствующие у других видов:
а) терминальные Q-сегменты: после окрашивания акрихин-ипритом на концах многих хромосом Pan и Gorilla (как правило, на одних и тех же хромосомных плечах) выявляются небольшие Q-сегменты. У Pongo так же, как и у Homo, они отсутствуют. Существуют два возможных объяснения данного факта: либо эти сегменты - результат мутации de novo у общего предка Pan и Gorilla, либо они имелись у примитивного предка современных гоминоидов и были утрачены в ходе эволюции Pongo и Homo. Обе гипотезы кажутся маловероятными, если исходить из классических представлений о дупликациях или делениях отдельных хромосомных сегментов как изолированных и случайных событиях;
б) гетерохроматиновые районы выявляются в коротком плече некоторых акроцентрических хромосом. Их число уменьшается в следующем ряду организмов в последовательности: Pongo → Gorilla → Homo → Pan. По-видимому, такой гетерохроматиновый материал время от времени образуется в виде мутаций de novo в непосредственной близости от центромер акроцентрических хромосом, а затем перемещается в результате случайных хромосомных перестроек в другие участки хромосом. Возможно, что подобный материал содержится во вторичной перетяжке 9-й хромосомы Homo (разд. 2.1.2). 9-е хромосомы человека и шимпанзе включают также гетерохроматиновый блок вблизи центромеры;
в) существуют вариации и в локализации Т-сегментов. Единственное возможное объяснение этого факта состоит в том, что периодически материал Т-сегментов на концах хромосомы синтезируется de novo и затем распределяется по другим хромосомным участкам в результате перестроек;
г) у горилл и человека выявлены дополнительные Q-сегменты вблизи центромер 3-й и 4-й хромосом; сходные сегменты обнаружены в хромосоме 9 гориллы, в хромосоме 13 человека и в 3-й хромосоме шимпанзе. У орангутана и гиббона их нет совсем.
Все эти три результата, полученные при изучении терминальных Q- и Т-сегментов, гетерохроматиновых районов и прицентромерных Q-областей, свидетельствуют о том, что кариотипические различия между пятью рассматриваемыми близкородственными видами обусловлены не только перестройками хромосом, которые можно объяснить, используя классические принципы. По-видимому, какую-то роль в этом играют дополнительные механизмы, например синтез de novo, а также потеря хромосомного материала. Исследования на уровне ДНК расширяют наше представление об этих процессах.
Недавно было построено гипотетическое филогенетическое древо приматов, отражающее взаимосвязи широкого круга таксонов от полуобезьян до человека. Оно основано на результатах изучения кариотипов более чем 60 видов этого отряда, проводившегося с использованием всех имеющихся методов дифференциального окрашивания [1913]. Эухроматиновый материал, т. е. невариабельные R- и Q-сегменты (разд. 2.1.2.1), у обезьян, человекообразных обезьян и у людей, по-видимому, идентичны. Все количественные и качественные изменения связаны с гетерохроматином. Типы хромосомных перестроек, выявляемые по видовым различиям в структуре хромосом, варьируют от подгруппы к подгруппе: например, у Lemuridae преобладают робертсоновские транслокации (центрические слияния); у Cercopithecinae часто встречаются хромосомные разрывы; для эволюции Pongidae, а также человека обычны перицентрические инверсии.
Хромосомные перестройки в ходе эволюции и в современной популяции. Имеется существенное различие между хромосомными перестройками, зафиксированными в ходе эволюции и возникающими в современной популяции. В настоящее время наиболее распространены центрические слияния хромосом D- и G-групп, сопряженные с утратой материала коротких плеч (разд. 2.2.2). Как это ни удивительно, в ходе эволюции понгид и человека ни одно, буквально ни одно из центрических слияний не зафиксировалось. Единственное объяснение этого факта состоит в том, что хромосомные перестройки такого рода селективно вредны, что может быть связано с образованием анеуплоидов, например зигот с трисомией по длинному плечу 21q (синдром Дауна) или нежизнеспособных анеуплоидных эмбрионов. Однако, насколько нам известно, не все центрические слияния оказываются селективно вредными. (Более подробно эти вопросы обсуждаются в разд. 2.2.2.2.)
Может быть высокая частота центрических слияний и потерь зигот, к которым они приводят, - это относительно недавно возникшая генетическая адаптация к специфическим особенностям размножения и развития людей?
Селективное преимущество высокой частоты спонтанных абортов у людей [1937]. Известно, что 5-7% всех зачатий у человека приводят к образованию эмбрионов с хромосомными аномалиями (разд. 2.2.4); большинство из них нежизнеспособны и абортируются. В редких случаях рождаются дети с тяжелыми уродствами, не имеющие шансов на выживание в примитивных жизненных условиях. У выживших индивидов, основная часть которых - анеуплоиды по Х-хромосоме, фертильность значительно снижена (разд. 2.2.3.2). Гибель зигот до их имплантации в большинстве случаев проходит незамеченной. На первый взгляд значительная потеря эмбрионов, обусловленная хромосомными аберрациями, свидетельствует о существенном понижении репродуктивной приспособленности вида, к которому мы принадлежим. Однако на эту проблему можно посмотреть с другой стороны. Известно, что у людей потомство на протяжении длительного времени нуждается в большой родительской заботе. Следовательно, между родами живых детей должен выдерживаться некий оптимальный интервал, максимизирующий вероятность того, что большая часть потомков доживет до репродуктивного возраста. Любой механизм, обусловливающий уменьшение числа рождений живых детей от максимального до оптимального и увеличение интервала между родами от кратчайшего до оптимального (не создающий при этом угрозы жизни матери), может давать селективное преимущество. Ранние выкидыши, обусловленные хромосомными аберрациями, при тех примитивных условиях, в которых существовали наши предки, вероятно, служили именно таким механизмом. Благодаря ему значительно увеличивалась средняя продолжительность грудного вскармливания, что, вероятно, уберегало детей от недоедания и кишечных инфекций. Высокая частота спонтанных абортов, очевидно, обусловливает уменьшение общего числа младенцев и малолетних детей, опекаемых одной матерью.
Относительно высокая частота центрических слияний может также быть связана с отбором. Этот механизм, вероятно, детерминирован ядрышковым организатором, так как участки хромосом, задействованные в процессах центрического слияния, концентрируются именно в этом районе. Из данной гипотезы следует, что у других высших приматов центрические слияния должны встречаться реже, чем у людей. Из-за малого размера популяций всех отличных от человека приматов, изучавшихся до настоящего времени, соответствующие данные о частоте центрических слияний - и хромосомных аберраций вообще - отсутствуют. С другой стороны, трисомии, являющиеся наиболее существенным источником репродуктивных потерь, обусловленных хромосомными аномалиями, у приматов, отличных от человека, несомненно имеются, о чем свидетельствует обнаружение трисомии по 21-й хромосоме у шимпанзе [1951]. Однако недостаток данных не позволяет прийти к каким-либо выводам относительно частоты этой аномалии; учитывая сравнительно малое число обследованных особей, резонно предположить, что у шимпанзе трисомия 21 встречается, возможно, не намного реже, чем у людей.
Гомология хромосом и хромосомных сегментов человека и сравнительно далеких от него видов, не принадлежащих к приматам. Гомологии в структуре хромосом и порядке генов можно обнаружить не только у различных приматов, включая человека, но и у видов, находящихся в более отдаленном родстве друг с другом. Например, локусы, сцепленные у человека, проявляют тенденцию к сцеплению и у мыши, наиболее хорошо изученного в генетическом отношении млекопитающего [1910; 1949]. Эти гомологии так сильны, что напрашивается вывод о сохранении различных групп сцепления у разных видов в результате действия естественного отбора. Такой вывод эквивалентен тезису о функциональном значении последовательности и порядка расположения генетического материала на уровнях более высоких, чем уровень отдельных генов (см. разд. 2.3 и 3.5.5). Х-хромосома оставалась почти неизменной на протяжении всей эволюции млекопитающих [156]; в Х-хромосомах мыши и человека обнаружены гомологичные группы, состоящие по крайней мере из десяти сцепленных локусов [1910; 1932]. Оно [156] рассматривал гипотезу, согласно которой причиной этого феномена могут быть инактивация Х-хромосом и дозовая компенсация.
Для генома в целом оценка средней длины хромосомных сегментов, сохранившихся со времени расхождения предков людей и мышей, полученная путем сравнения двух карт сцепления, составляет ≈ 8,1 см [1960]. Эта оценка была использована для определения числа хромосомных перестроек, зафиксированных в двух предковых популяциях за время, прошедшее после их разделения.
Как может осуществляться фиксация хромосомной перестройки в популяции? Как показано в разд. 6.4.2, подавляющее большинство новых мутаций, возникающих в популяции, утрачивается. Такая потеря происходит не только в случае селективно нейтральных, но и в случае селективно ценных мутаций. Большинство хромосомных перестроек, как, например, перицентрические инверсии, часто оказываются селективно вредными, поскольку приводят к нарушениям мейоза. С другой стороны, Кимура (1968) [1510] показал, что скорость фиксации почти нейтральной мутации зависит только от соответствующей частоты мутирования. Частоты мутаций, приводящих к возникновению перицентрических инверсий, пока неизвестны. Не зная их, а также не имея достоверных данных о степени селективной вредности, которая может варьировать для разных перестроек, выдвигать какие-либо гипотезы относительно вероятности фиксации невозможно.
Кроме того, мы должны учитывать одну специфическую особенность отбора против инверсий. Такой отбор действует только против гетерозигот. Гомозиготы по инверсиям имеют нормальную фертильность независимо от локализации инверсии, поскольку спаривание гомологичных хромосом в мейозе протекает уже нормально. Известна ли нам какая-либо генетическая ситуация, в которой происходит быстрый выход из "опасного" состояния гетерозиготности, скажем, всего за два поколения? Такая ситуация может создаваться, когда брат и сестра наследуют от одного из своих родителей одну и ту же перестройку и производят в инцестном браке гомозиготное потомство. В этой группе гомозигот фертильность опять была бы нормальной, тогда как скрещивания в общей популяции дали бы только гетерозиготных потомков, имеющих пониженную плодовитость. Следовательно, этот механизм воздвиг бы эффективный репродуктивный барьер, что создало бы наилучшие условия для постепенного становления нового вида (рис. 7.6) [1947].
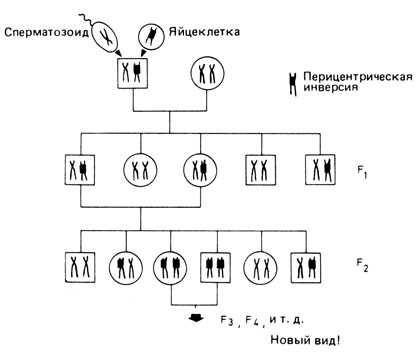
Рис. 7.6. Возможный способ образования нового вида в процессе тесного инбридинга. В половой клетке некоторой особи возникает перицентрическая инверсия, что приводит к появлению в следующем поколении одного гетерозиготного носителя. Этот носитель может иметь несколько гетерозиготных потомков, которые будут скрещиваться друг с другом и давать гомозиготное потомство
Современные приматы часто живут небольшими группами. Такой образ жизни, вероятно, вели и наши предки предлюди. Это увеличивало вероятность близкородственных скрещиваний. Однако если уже в этих предковых группах действовало "табу инцеста", предотвращающее материнско-сыновние или брато-сестринские браки, то до появления двух гомозигот, образовавших предковую пару, могло пройти одно или несколько большее число поколений гетерозигот. Отметим, что гомозиготность по перицентрической инверсии была обнаружена в современной человеческой популяции у ребенка, родившегося в браке отца с дочерью [1902]. Может ли быть так, что новые виды приматов берут начало от одной пары особей? Или более конкретно: реально ли, что все человеческие существа происходят от одной предковой пары? Как это ни удивительно, миф об Адаме и Еве как паре прародителей человечества может со временем получить научное обоснование. Сравнительное изучение 1511 видов, представляющих 225 родов позвоночных, выявило сильную корреляцию между скоростью хромосомной эволюции и видообразованием и показало, что оба этих процесса протекают у приматов очень быстро [1905]. Авторы данной работы представили доказательства того, что решающим фактором, влияющим на эти процессы, является, вероятно, подразделение популяций на небольшие демы. Этот вывод полностью согласуется с обсуждавшейся выше гипотезой.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'