8.2.3. Новые подходы, предложенные для исследования генетики поведения человека
Большая часть материала, охватываемого в разд. 8.2.3, относится к возможному применению генетических понятий к поведенческим явлениям. Основные генетические понятия были раскрыты в предшествующих главах, особенно в гл. 3 и 4. Для анализа проблем генетики поведения до сих пор они не использовались в полную силу. Вполне возможно, однако, что для удовлетворительного объяснения генетической детерминированности поведения человека этих понятий окажется недостаточно и мы не сумеем это сделать так же успешно, как, например, в случае серповидноклеточной анемии.
Во введении к этой книге мы говорили о том, что генетический подход к биологическим явлениям является "редукционистским": генетический анализ считается успешным, если наследуемое различие можно проследить до различия в состоянии гена. В большинстве случаев - например, когда мы хотим понять природу недостаточности фермента, этот подход приводит к удовлетворительным результатам. Его ограничения становятся очевидными, когда мы пытаемся анализировать генетическую изменчивость при нормальном эмбриональном развитии или отклонения, которые ведут к врожденным порокам развития. В этой ситуации сложная система механизмов обратной связи, по-видимому, регулирует активность генов в различных группах клеток и в разные фазы развития. Развитие поведения человека не заканчивается никогда, оно продолжается в течение всей его жизни. Включенные в него механизмы обратной связи, вероятно, более сложны, чем механизмы, действующие во время эмбрионального соматического развития. Возможно, что для некоторых психических функций в ходе эволюции появились новые принципы действия генов. Таким образом, современные генетические подходы, которые оказались столь плодотворными в других областях, при анализе генетики поведения могут не принести успеха. И все-таки мы должны попробовать применить их, поскольку это единственный способ выявить их ограничения. Не исключено, что в будущем возникнут новые идеи, с помощью которых удастся преодолеть теперешние трудности.
8.2.3.1. Генетическая изменчивость, которая может влиять на поведение человека
В настоящее время данных о генетической изменчивости, влияющей на поведение человека, мало. Мы довольно детально описываем возможные подходы к изучению этой проблемы потому, что считаем необходимым дальнейшее ее развитие [1968] (рис. 8.21).
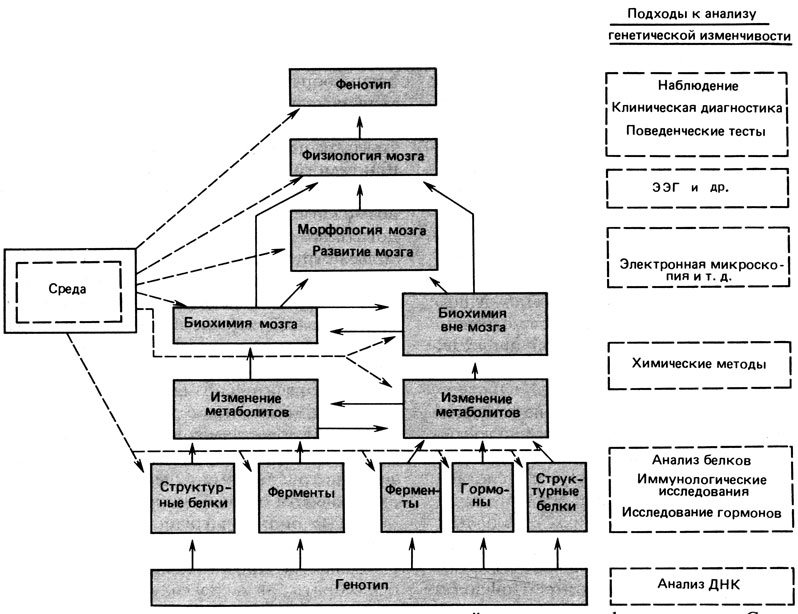
Рис. 8.21. Возможные уровни исследования генетической изменчивости функции мозга. Сплошной контур - уровень, на котором может наблюдаться генетическая изменчивость; пунктирный контур - метод исследования
Общий метаболизм. Грубые нарушения метаболизма приводят к затемнению сознания и остановке психических процессов. Примерами таких нарушений, не обусловленных генетически, являются недостаточность функции печени, приводящая к печеночной коме, и почечная недостаточность, вызывающая уремию. Механизмы развития обоих состояний представляются ясными по крайней мере в общих чертах. Организм теряет способность правильно метаболизировать или выделять определенные соединения, их концентрация нарастает, они взаимодействуют с метаболитами мозга, нарушая идущие в норме процессы, и если эти нарушения достаточно тяжелы, то развивающаяся в итоге интоксикация уничтожает психическую деятельность.
Интоксикация может изменять функции мозга не только когда токсическое вещество образуется в процессе метаболизма в организме, но и тогда, когда оно поступает извне. Изменения сознания и восприятия, которые наступают после приема пищи, содержащей природные яды, наблюдались довольно рано в истории развития человеческого рода. В современном обществе наркомания представляет собой грозную социальную проблему. С другой стороны, сегодняшняя медицина научилась широко использовать действие веществ, влияющих на нервную систему: от общей анестезии до лечения больных психическими заболеваниями. Прежде чем проникнуть в мозг и изменить его работу, вещества подвергаются метаболизму, изменяющему их химический состав и модифицирующему их действие. Эти метаболические изменения опосредованы главным образом различными ферментами. Ферменты в свою очередь могут демонстрировать генетическую изменчивость, которая влияет на их активность, специфичность к субстрату и другие характеристики (разд. 4.2). Вытекающие отсюда различия в действии веществ, а также в побочных эффектах являются предметом исследования фармакогенетики (разд. 4.5.1). Генетическая изменчивость характерна для ферментов, метаболизирующих соединения, содержащиеся в потребляемой нами нормальной пище и необходимые для определенных метаболических процессов. В некоторых случаях генетическое изменение молекулы фермента может быть столь основательным, что его активность полностью или почти полностью исчезает. В качестве примера такого изменения, затрагивающего развитие и функции мозга, можно привести недостаток фенилаланингидроксилазы при фенилкетонурии, который приводит к "наводнению" мозга фенилаланином и другими токсичными метаболитами. Другой пример - состояние хронической аммиачной интоксикации, обусловленное ферментативными нарушениями в цикле мочевины. Многие врожденные ошибки метаболизма связаны с умственной отсталостью, детерминируемой аналогичными механизмами.
Изменчивость гормонов. Многие процессы в организме человека опосредованы гормонами (разд. 4.7.). Известно большое число количественных и качественных генетических дефектов гормональных функций. Большинство из них затрагивает самочувствие человека и его поведение. Наглядным примером является недостаточность функции щитовидной железы, которая приводит к гипотиреозу с типичным для него отсутствием жизненного тонуса. Половые гормоны также обладают сильным действием на эмбриональное и постнатальное развитие, на поведение и психическое состояние взрослых людей. При поиске генетических механизмов, которые могли бы влиять на поведение человека, эндокринные железы будут, следовательно, основным кандидатом на проверку их генетической изменчивости.
Генетическая изменчивость внутри мозга. Генетические различия в количестве производимых гормонов или в их молекулярной структуре являются только одним из возможных источников генетической изменчивости гормональной функции. Второй источник такой изменчивости - действие гормонов на органы-мишени. В этом процессе важными посредниками являются рецепторы гормонов. Некоторые генетически обусловленные дефекты, которые в настоящее время классифицируют как рецепторные заболевания (разд. 4.6.4), затрагивают функции мозга. Существует гипотеза, согласно которой генетическая изменчивость гормональных рецепторов в мозге может влиять на функции мозга и служить, таким образом, причиной генетической изменчивости поведения.
Это только один пример того, как генетическая изменчивость структуры и функции самого мозга может влиять на изменчивость поведения. Чтобы раскрыть другие возможные механизмы, мы должны обследовать все уровни, на которых может изучаться структура и функция мозга - от анатомии и гистологии до электрофизиологии и основных биохимических процессов, включенных в возбуждение и торможение нервных клеток. Во всех этих областях накоплено гигантское количество информации, однако поразительно мало сделано для того, чтобы "просеять" эту информацию для выявления генетической изменчивости. В какой-то степени это объясняется методическими трудностями. Оценка генетической изменчивости обычно требует исследования довольно больших групп индивидов. Для признаков крови это вполне реально.
Если бы пробы ткани мозга можно было брать так же легко, как пробы крови, наши знания о генетической изменчивости, которая влияет на поведение человека, были бы полнее. Следует, однако, отдавать себе отчет в том, что даже успехи в изучении генетического полиморфизма, влияющего на мозг, вряд ли позволят объяснить поведение человека, поскольку детерминанты поведения гораздо сложнее, чем, например, функции эритроцитов. Вот почему необходимы разнообразные подходы.
1. Один из них связан с поиском генетической изменчивости в физиологических характеристиках, которые можно измерять прямо с помощью неинвазивных методов. Наибольшие возможности до сих пор предоставлял генетический анализ нормальной электроэнцефалограммы (ЭЭГ) [921]. Недостатком этого подхода является непрямая связь между действием гена и физиологическим фенотипом. Она, несомненно, больше подвержена возмущающим воздействиям со стороны промежуточных переменных, чем, например, фермент.
2. Можно искать генетическую изменчивость, которая выражается не только в мозговой, но и в других тканях (лучше всего в различных типах клеток крови, в которых может выявляться действие генов, активных в мозге). Ограничения этого подхода связаны с биохимической дифференцировкой во время эмбрионального развития, которая уменьшает возможность экстраполяции данных, полученных на одной ткани, на другую. Есть основания полагать, что дифференцировка и функциональная специализация могут быть более всего развиты в центральной нервной системе, что делает экстраполяции с других тканей особенно рискованными. Все же этот подход дал ряд многообещающих результатов при исследовании, например, тромбоцитов и ферментов сыворотки крови у больных аффективными психозами. В некоторых случаях гены, не проявляющие своего действия в доступной для анализа ткани, в других тканевых системах могут быть даже активированы.
3. Генетическую изменчивость и ее связь с поведением можно исследовать в мозге экспериментальных животных. Этот подход опирается на известную гомологию между человеком и другими млекопитающими во многих физиологических процессах, которая основана на их общем происхождении в филогенезе. Животные как модельная система широко используются для генетического анализа в ситуации, когда исследование на человеке не может быть проведено по этическим причинам, например при исследовании индуцированного мутагенеза [2142]. Очевидным преимуществом этого подхода для генетики поведения является прямой доступ к мозгу. Его недостаток определяется существованием межвидовых различий и уникальной ролью человеческого мозга. Данный подход, таким образом, помогает сформулировать модели механизмов, с помощью которых различия в физиологии мозга приводят к различиям в поведении. В процессе анализа таких моделей могут возникнуть идеи о том, где локализуются соответствующие различия. При дальнейшем обсуждении вопроса мы будем опираться главным образом на результаты экспериментов с животными.
Популяции экспериментальных животных обычно формируются из инбредных линий и по структуре воспроизводства они радикально отличаются от человеческих популяций. Животные инбредных линий генетически идентичны. Их можно сравнить с монозиготными близнецами у людей. Результаты, полученные на одной инбредной линии, следует сопоставлять с результатами, полученными на других линиях и даже в естественных популяциях.
Перечисленные подходы к изучению генетической изменчивости мозговых функций корректны, если принципиальные положения генетики, т. е. понятие гена, регуляции действия гена, генетически определяемых ферментов и рецепторов, справедливы для мозговых функций. Вполне возможно, что понимание сложного, разумного поведения потребует дополнительных знаний об основах обработки и организации информации. На рис. 8.21 дана общая схема тех уровней, на которых генетическая изменчивость функций мозга может вызывать соответствующую изменчивость поведения.
8.2.3.2. Генетическая изменчивость вне мозга, влияющая на поведение человека
Дефекты ферментных систем, которые приводят к умственной отсталости (рис. 8.22). Многие из известных дефектов ферментных систем у человека приводят к умственной отсталости и другим аномальным проявлениям. Человеческий мозг особенно чувствителен к разного рода нарушениям метаболизма (табл. 8.15), однако для понимания нормальной работы мозга эти наблюдения до сих пор дали очень мало. Вполне вероятно, например, что нервные клетки не могут полноценно работать, если они "перенаселены" промежуточными метаболитами типа тех, которые обнаружены при мукополисахаридозах [1240] или муколипидозах. По-видимому, развитие и деятельность мозга особенно чувствительны к изменениям биохимической среды. Врожденные ошибки метаболизма могут приводить к увеличению или снижению содержания множества веществ, что часто служит причиной умственной отсталости. Сопоставление наследственных отклонений, не вызывающих патологию психики, с теми, которые ее вызывают, хотя и дает в руки исследователей кое-какую информацию, но не объясняет нормальную работу мозга.
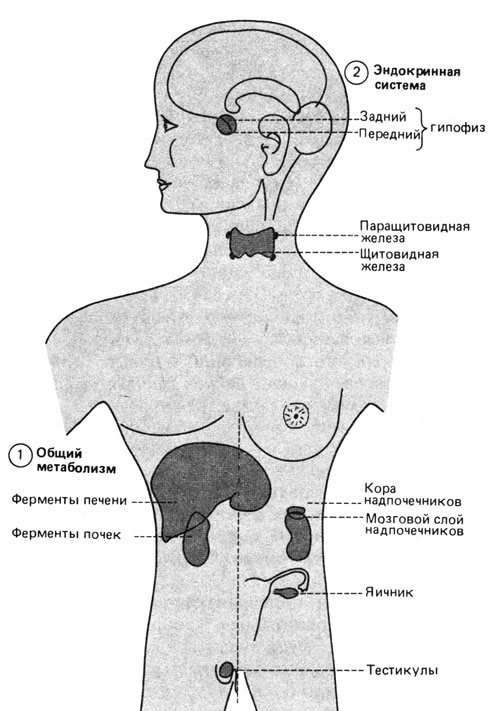
Рис. 8.22. Генетическая изменчивость вне мозга, которая может оказывать влияние на поведение человека. В общем плане наиболее существенны метаболические влияния ферментов печени и почек. Источником других воздействий могут быть железы внутренней секреции, такие, как передний и задний отделы гипофиза, надпочечники, половые железы, щитовидная и паращитовидная железы

Таблица 8.15. Отдельные типы врожденных нарушений метаболизма, ведущих к умственной отсталости
Генетически определяемые аномалии метаболизма могут приводить не только к умственной отсталости, в некоторых случаях они вносят свой вклад в подверженность таким психическим заболеваниям, которые объединяются понятием "шизофрения".
Членовредительство при синдроме Леша - Найхана: мочевая кислота. Синдром Леша - Найхана был описан в разд. 4.2.2.6 как синдром, вызванный недостаточностью фермента гипоксантин-гуанин - фосфорибозилтрансферазы. Этот фермент необходим для использования гипоксантина в синтезе гуанина. При его недостатке большое количество гипоксантина превращается в мочевую кислоту. Больные страдают повышенной возбудимостью, приводящей к гиперрефлексии, для них характерны почти постоянно совершаемые движения, а также компульсивное стремление к членовредительству. Несмотря на боль, они кусают свои пальцы и губы и калечат себя. Эта склонность к самокалечению не имеет никаких аналогий в нормальной или патологической психологии человека. Нам известны и другие типы компульсивных неврозов. Некоторые больные, например, характеризуются навязчивым стремлением без конца мыть руки. Среди родственников больных компульсивными неврозами шизофреноподобные психозы, по-видимому, встречаются чаще, чем в общей популяции [2187]. Возможно, анализ специфического повреждения, вызываемого недостатком гипоксантин-гуанин-фосфорибозилтрансферазы в мозге, мог бы дать кое-какие сведения относительно нейрональных механизмов импульсивного поведения. Кроме того, было бы интересно выяснить, имеются ли у гетерозиготных по этому дефекту индивидов какие-либо странности в поведении, особенно когда они достигают среднего или пожилого возраста.
Давно описана и неоднократно подтверждалась положительная корреляция между уровнем мочевой кислоты в крови и IQ. Многие исторические личности страдали подагрой. Можно предположить, что мочевая кислота немного увеличивает возбудимость нейронов и эта неспецифическая стимуляция положительно влияет на интеллект. Однако проведенное недавно Проппингом исследование на близнецах не обнаружило корреляции между уровнем мочевой кислоты и интеллектом.
Гетерозиготны по рецессивным заболеваниям. Активность ферментов у лиц, гетерозиготных по врожденным расстройствам метаболизма, обычно вдвое ниже нормальной. Поэтому неудивительно выявление фенотипических нарушений, по крайней мере в случае, если специфические метаболические пути находятся в постоянном напряжении или по мере старения организма. Однако систематические исследования таких гетерозигот, особенно исследования их психического состояния, чрезвычайно малочисленны (см. также разд. 4.2.2.8). Имеющиеся в литературе данные недавно проанализированы одним из нас в обзорной статье [1340]. Наиболее обстоятельные исследования были выполнены на больных фенилкетонурией (ФКУ). Несмотря на недостатки эпидемиологических методов, использовавшихся в некоторых из этих работ, можно сделать ряд выводов.
1. Гетерозиготы по ФКУ имеют более низкие показатели по ряду пунктов IQ в сравнении с показателями контрольной группы. Показатели по вербальному набору теста снижаются, по-видимому, в большей степени, чем по тестам праксиса. 2. Некоторые исследования указывают на несколько повышенный риск психотических расстройств и психозов, имеющих позднее начало, злокачественное течение и включающих симптомы депрессии. 3. У некоторых гетерозигот по данным ЭЭГ обнаружен повышенный уровень возбудимости коры. 4. Эти небольшие аномалии могли быть вызваны (по крайней мере частично) увеличением уровней внутриклеточного фенилаланина и тирозина, поскольку было обнаружено, что содержание этих аминокислот увеличено в лимфоцитах (более доступные клетки).
Другие аутосомные рецессивные расстройства, при которых были описаны небольшие фенотипические отклонения, включают несколько форм липидозов, таких, как метахроматическая лейкодистрофия позднего детского возраста (25010), глобоидно-клеточная лейкодистрофия Краббе (24520). Отклонения обнаружены и у гетерозигот по генам болезни Зандхоффа (26880), болезни Нимана-Пика (25720) и болезни Вольмана (27800). В этих случаях небольшие отклонения выявлены главным образом в тестах IQ, связанных с выполнением действий, особенно в тестах на пространственное восприятие. Кроме того, были увеличены показатели по личностным опросникам, тестирующим психосоматические расстройства, депрессию и эмоциональную лабильность. И снова причиной отклонений могло быть накопление аномальных метаболитов, так как некоторое увеличение содержания муколипидов обнаруживается не только у гомозигот, но и у гетерозигот (рис. 8.23).
![Рис. 8.23. Лимфоциты гетерозигот по юношеской амавротической идиотии. В клетках выявляются похожие на вакуоли структуры с липидными включениями [1060]](pic/000060.jpg)
Рис. 8.23. Лимфоциты гетерозигот по юношеской амавротической идиотии. В клетках выявляются похожие на вакуоли структуры с липидными включениями [1060]
В двух исследованиях аутосомных рецессивных микроцефалии (25120), проведенных в разных частях света (в Канаде и Советском Союзе), было обнаружено, что около одной трети гетерозигот с нормальным мозгом имеют сниженный интеллект. По мнению Квази и Рида [2172], авторов работы, проведенной в Канаде, такие гетерозиготы могут составлять ощутимую долю всех лиц, страдающих умственной отсталостью "неизвестной природы". По их представлению гомозиготы встречаются с частотой 1:40000, а частота гетерозигот примерно 1:100 (разд. 3.2.1). Если одна треть из них страдает умственной отсталостью, это означает, что примерно одна треть от 1% всего населения является умственно отсталой, потому что несет один ген микроцефалии. Распространенность умственной отсталости в Соединенных Штатах и Англии при использовании в качестве порога величины IQ, равной 69, была оценена приблизительно в три процента [2157]. Таким образом, один из девяти умственно отсталых индивидов из общей популяции может быть гетерозиготным по гену микроцефалии. Конечно, эта оценка сугубо приблизительна; в других популяциях рецессивная микроцефалия может быть более редкой, и, кроме того, возможно существование генетической гетерогенности. Важно, однако, что в принципе этот вывод вполне реален. Его можно сформулировать таким образом: если одна треть таких гетерозигот страдает умственной отсталостью, другие две трети, вероятно, будут иметь IQ у нижней границы нормы. Кроме того, примерно 1 из 50 членов нашей популяции гетерозиготен по ФКУ (принимая частоту гомозигот приблизительно за 1:10000). Небольшое снижение IQ описано и для гетерозигот по другим заболеваниям. Следует помнить, что многие формы умственной отсталости, связанной с аутосомно-рецессивными или Х-хромосомными генами, пока неполностью охарактеризованы. Если учесть также, что довольно распространенные формы умственной неполноценности, связанные с Х-хромосомой, могут выражаться не в отсталости в полном смысле, а в снижении IQ, то можно утверждать, что значительная часть "нормальной" генетической изменчивости IQ в области низких значений может быть обусловлена гетерозиготностью по аутосомным и сцепленным с Х-хромосомой рецессивным заболеваниям.
8.2.3.3. Действие гормонов
Как действуют гормоны! Известно, что гормоны обычно действуют на специальные клетки, которые имеют рецепторы, связывающие гормоны. Это запускает в клетках синтез специфических белков; роль посредника в данном процессе играет циклический AMP. У человека описаны некоторые "рецепторные расстройства"; наиболее известные из них - семейная гиперхолестеринемия и тестикулярная феминизация (разд. 4.7.5). В последнем случае нечувствительность рецепторов к андрогенам приводит к развитию женского фенотипа у индивидов, имеющих XY-кариотип и, следовательно, семенники.
Рецепторы к гормонам присутствуют и на клетках ЦНС; на их развитие и функции могут оказать влияние по крайней мере три фактора: метаболические процессы, вызванные гормонами в других тканях, которые влияют на функции мозга непрямым способом, аномальное снабжение гормонами в количественном или качественном отношении, индивидуальные различия в рецепторах. Например, действие тироксина на психическую и нервную активность (которое производит большое впечатление, когда видишь, как больной гипотиреозом отвечает на гормональную терапию) является, вероятно, непрямым, так как тироксин увеличивает базальную скорость метаболизма во всех тканях, кроме мозговой [120].
С другой стороны, половые гормоны, по-видимому, прямо влияют на развитие мозга [138]. Это влияние начинается уже в эмбриональном периоде, о чем свидетельствуют наблюдения над девочками, которые подверглись маскулинизации вследствие введения матерям синтетических прогестинов.
Томбойизм (мальчишеские Повадки) у девочек, подвергавшихся действию маскулинизирующих веществ в пренатальный период [138]. Томбойизм можно охарактеризовать следующим образом.
1. Эти девочки любят играть с мальчиками в спортивные игры, особенно в игры с мячом, и предпочитают те же игрушки, что и мальчики.
2. Их уверенность в себе и настойчивость в борьбе за лидерство достаточно высоки для того, чтобы успешно конкурировать с мальчиками. При этом они не агрессивны.
3. Эти девочки обычно предпочитают носить брюки и шорты, хотя в особых случаях и не прочь одеться по-женски.
4. Они, как правило, не играют в куклы, не любят няньчиться с детьми.
5. Желание иметь дружка появляется у такой девочки позже, чем у большинства ее ровесниц. Любовь и замужество находятся на втором плане по сравнению с личными достижениями и карьерой. У них, однако, нет никакой склонности к лесбианству.
Получены данные о том, что томбойизм может быть обусловлен действием гормонов в период эмбрионального развития. Прогестины - это стероиды, родственные по химической структуре андрогенам. В функции сохранения беременности они могут замещать прогестерон. Когда прогестины впервые использовали в терапевтических целях, маскулинизирующее действие этих гормонов не было известно. В 1950-х годах у некоторых женщин родились дочери, нормальные во всех отношениях, за исключением маскулинизации клитора. Маскулинизирующее действие гормона прекращалось в момент рождения, и такие дети развивались и созревали как девочки. К другой группе девочек с признаками маскулинизации относятся больные с адреногенитальным синдромом, обусловленным аутосомно-рецессивным дефектом 21-гидроксилазы - одного из ферментов, необходимых для синтеза кортизола (разд. 4.2). Это состояние корректируется с помощью введения кортизола, который, снижая выделение гипофизом АКТГ, резко уменьшает синтез предшественников кортизола, имеющих андрогеноподобное действие.
В табл. 8.16 сопоставлены характеристики поведения дочерей тех женщин, которым вводили прогестин во время беременности, пациенток с адреногенитальным синдромом с соответствующими характеристиками пациенток с тестикулярной феминизацией и с синдромом Тернера. Последние не подвергаются действию половых гормонов в период эмбрионального развития, так как у них нет гонад. В момент исследования самой старшей девочке из группы, подвергшейся действию прогестина, было 16 лет; возраст пациенток с синдромом Тернера колебался в пределах от 8 до 16,5 лет. Томбойизм в той форме, как описано выше, был выявлен у 9 из 10 девочек, испытавших действие прогестина, у 11 из 15 девочек с адреногенитальным синдромом, но практически отсутствовал у пациенток с синдромом Тернера и с тестикулярной феминизацией. Соответствующим образом было модифицировано отношение всех этих девочек к их будущей роли замужней женщины и матери.
![Таблица 8.16. Томбойизм (мальчишеское поведение) среди девочек с разными генетическими аномалиями [138]](pic/000242.jpg)
Таблица 8.16. Томбойизм (мальчишеское поведение) среди девочек с разными генетическими аномалиями [138]
Полученные результаты подтверждались данными экспериментов на самках макаков-резусов, которых искусственно андрогенизировали в период эмбрионального развития и которые в детском периоде демонстрировали поведение, сходное с поведением самцов того же возраста [138]. Все это говорит о том, что специфичные для каждого пола системы мозга оказывают влияние на формирование отношения детей и подростков к выполнению своей роли в качестве представителя определенного пола.
Тестикулярная феминизация. Синдром тестикулярной феминизации (31370) обусловлен нечувствительностью к андрогенам. Если в мозге тоже отсутствуют функциональные рецепторы к андрогенам, можно ожидать от таких индивидов "типично женского" отношения к браку и материнству. Оно действительно было выявлено у большинства пациенток, которых удалось обследовать [138]. Однако интерпретация результатов здесь не столь однозначная, как в случае девочек, подвергавшихся пренатальным маскулинизирующим воздействиям. Пациенты с тестикулярной феминизацией почти ничем не отличаются по внешнему виду от девочек и также развиваются; следовательно, их идентификацию с женской ролью можно объяснить с чисто психологических позиций.
После появления этих сообщений вопрос неоднократно исследовали на животных и человеке, но результаты не всегда были однозначными (см., например, [2030; 2128]). Однако при знакомстве с литературой у нас сложилось впечатление, что пренатальное воздействие андрогенами все-таки влияет на развитие мозга.
Гомосексуальность и гормоны. Исследования на близнецах, рассматривавшиеся в разд. 8.2.1.5, указывали на генетическую компоненту в гомосексуальном поведении мужчин. Предполагалось, что искажение схемы сексуальной разрядки у мужчин-гомосексуалистов может быть вызвано патологически низким содержанием андрогенов. Чтобы проверить это, измеряли уровень стероидных гормонов в моче и сыворотке крови [138]. Выяснилось, что соотношение определенных метаболитов половых гормонов, обнаруживаемых в моче, отличается от нормального; однако в этих исследованиях недостаточно тщательно были учтены другие факторы - возраст, степень половой активности и общее состояние здоровья. Кроме того, колебания уровня этих метаболитов в моче контрольных лиц довольно значительны, и диапазон колебаний перекрывает те изменения, которые обнаруживаются у гомосексуалистов.
Даже мужчины, которым предстояло лечение эстрогенами и хирургическое вмешательство по половым показаниям, имели нормальный уровень андрогенов [2028].
Эти данные демонстрируют отсутствие прямой зависимости между выработкой андрогенов и гомосексуальностью. Но они не исключают возможной связи гомосексуальности с небольшими различиями в рецепторах к андрогенам. Чтобы подтвердить или отвергнуть эту гипотезу, еще предстоит поработать. Новые и независимые данные о половых различиях в функциях мозга получены в исследованиях, в которых применяется неинвазивный метод, позволяющий следить за функциональным состоянием центральной нервной системы - электроэнцефалограмма (ЭЭГ).
8.2.3.4. Физиология мозга: генетика ЭЭГ
Прямое исследование генетической изменчивости работы мозга связано с методическими трудностями. Вот почему необходимы косвенные методы. Некоторые из них были уже упомянуты: прямое изучение физиологии мозга, исследование генетической изменчивости вне мозга, потенциально связанной с его работой, эксперименты на животных. При изучении физиологии мозга использовалась главным образом электроэнцефалограмма (ЭЭГ).
ЭЭГ человека. Наиболее важные характеристики ЭЭГ человека были описаны в разд. 3.6.1, где на примере низковольтной ЭЭГ обсуждались критерии простого аутосомно-доминантного наследования количественного признака с непрерывной изменчивостью. Речь шла о том, что ЭЭГ покоя, имеющая в детстве нерегулярный характер с относительно медленными волнами, развивается в течение детского и юношеского периода и приобретает законченный, стабильный характер к 19 годам. В ЭЭГ взрослого человека преобладают α-волны (частота 8-13/с) с различной долей β-волн (≥ 14/с) и θ-волн (4-7/с); α-волны обычно в наибольшей степени выражены в затылочной области мозга. В нормальных условиях у здоровых людей различия между отведениями, сделанными в разное время, невелики у одного и того же человека; различия между разными людьми - весьма существенны.
Исследования на близнецах. ЭЭГ представляет собой очень сложный признак со многими переменными, в числе которых распределение частот и амплитуд в одном отведении, колебания между отведениями от различных областей головы и форма волн. Чтобы узнать, какова роль генетических факторов в этой изменчивости, разумно сравнить МЗ и ДЗ близнецов. Поскольку ЭЭГ по мере взросления меняется, самыми подходящими пробандами были близнецы в период первого и второго десятилетий их жизни и несколько молодых совершеннолетних лиц. Оказалось, что в отсутствие тяжелой усталости, болезней мозга типа эпилепсии или опухоли или тяжелой патологии метаболизма характер мозговых волн в стандартных условиях (расслабленное состояние с закрытыми глазами) практически полностью определяется генетически. Это заключение справедливо также для скорости созревания мозга, о которой свидетельствует развитие ЭЭГ [2228].
Можно было бы предположить, что причина конкордантности заключается в общей среде развития близнецов. Однако исследование ЭЭГ у 8 пар взрослых близнецов из Дании, которые воспитывались раздельно, выявило такую же степень конкордантности [2095]. Сильное сходство в ЭЭГ сохраняется до старости. Дискордантными могут быть только те аномалии в ЭЭГ, которые вызваны болезнью типа небольшого инсульта [2074]. Характер взрослой ЭЭГ идентичен даже у близнецовых пар с довольно разными эмоциональными переживаниями в прошлом, например когда один из близнецов страдал тяжелым неврозом. С другой стороны, органические заболевания мозга, такие, как эпилепсия, могут быть причиной четких и постоянных различий в ЭЭГ даже между МЗ близнецами.
Как уже упоминалось, развитие ЭЭГ в детском возрасте обнаруживает выраженную межиндивидуальную вариабельность и определяется, по данным близнецовых исследований, генетическими факторами. Психологическое созревание у разных индивидов тоже протекает с разной скоростью. Кроме того, у детей, которых психологи относят к незрелым или обнаруживающим аномалии поведения, часто наблюдался нерегулярный характер ЭЭГ. Данные такого рода позволяют предположить, что характер созревания ЭЭГ может быть связан с различиями психологического созревания в пределах нормы, которые определяются с помощью специальных тестов. Это предположение подтвердилось для α-частоты ЭЭГ [2233]. Следовательно, частота ЭЭГ служит мерой части генетической изменчивости, влияющей на индивидуальные различия в нормальном психологическом развитии. Эта связь заслуживает более детального изучения.
Семейные исследования. Результаты исследований на близнецах стимулировали поиск более четких свидетельств в пользу генетических механизмов. Если предполагается моногенный тип наследования, для анализа следует взять альтернативные признаки. Такие признаки действительно существуют, и наиболее явный из них - это так называемая низковольтная ЭЭГ с маловыраженной или отсутствующей α-активностью в затылочной области. Характер наследования оказался аутосомно-доминантным (разд. 3.6.1; рис. 3.54) со слабым перекрыванием между двумя классами фенотипов.
В другом наследственном типе ЭЭГ преобладают мономорфные α-волны. Средний α-ритм имеет максимум в затылочной области коры головного мозга и становится более нерегулярным и смешанным с другими волнами в передних отделах. При мономорфном типе α-волн очень регулярные α-волны высокой амплитуды обычно регистрируются по всей коре головного мозга. Близнецовые и семейные исследования не оставляют никаких сомнений в том, что этот тип является наследственным так же, как и низкоамплитудная ЭЭГ. Семейные данные указывают на простое доминирование [921]. Однако отграничить этот тип от средней ЭЭГ труднее, чем низкоамплитудную ЭЭГ.
В определенном смысле этот тип ЭЭГ можно рассматривать как "контртип" низкоамплитудной ЭЭГ; в то время как последняя имеет слабый α-ритм, в мономорфной α-ЭЭГ этот ритм представляется особенно сильным.
Существуют и другие признаки с простым доминантным наследованием - например, варианты, в которых затылочные α-волны замещены волнами, имеющими частоту 16-19/с и общие с α-волнами свойства (блокада при открывании глаз и при других видах стимуляции). В других семьях были обнаружены генетические варианты быстрого компонента - β-волн. В отличие от α-волн β-волны в большинстве случаев концентрируются во фронтальной и прецентральной областях коры; в некоторых семьях эти β-волны образуют характерные веретенообразные группы. Различают два аутосомно-доминантных типа [921].
Половые различия в структуре ЭЭГ. Большинство видов ЭЭГ с β-волнами обнаруживает выраженное накопление в определенных семьях; эти данные не согласуются с моделью простого моногенного наследования. Проще всего их объяснить с помощью мультифакториального наследования в сочетании с пороговыми эффектами (разд. 3.6.2). Кроме того, распространенность ЭЭГ с β-волнами (главным образом диффузными) увеличивается с возрастом, и по этому признаку существуют определенные половые различия. Во всех возрастных группах, за исключением детской, распространенность ЭЭГ с быстрыми волнами среди женщин выше, чем среди мужчин. Это различие между полами часто связано с немногочисленностью α-волн в затылочном отведении [2043].
Как мозг генерирует ЭЭП Наследственные разновидности ЭЭГ указывают на различия в физиологических функциях человеческого мозга. Для понимания природы этих различий и их возможного влияния на поведение, необходимо знать, каким образом генерируется ЭЭГ. В этом отношении данные экспериментальной нейрофизиологии рисуют относительно ясную картину [1998]. Волны ЭЭГ, особенно α-волны, образуются благодаря взаимодействию нейрофизиологических процессов на нескольких, по крайней мере трех-четырех, уровнях (рис. 8.24). "Батарея" ЭЭГ размещена в коре больших полушарий; расположенные здесь группы нейронов разряжаются в определенном ритме. Их активность, однако, координируется водителем ритма (точнее, группой связанных между собой водителей), расположенным в таламусе. Активность таламуса в свою очередь находится под влиянием входов от структур мозга, расположенных на более низких уровнях. Восходящая ретикулярная активирующая система (ВРАС) локализована в ретикулярной формации, главным образом в области варолиевого моста и продолговатого мозга. ВРАС играет ведущую роль в организации, например, сна и сновидений. В состоянии бодрствования она поддерживает определенный уровень "тонической активации", на который оказывают влияние входы от специфических, афферентных путей, возбуждаемых сенсорной стимуляцией. Высокий уровень активации вызывает десинхронизацию ЭЭГ. На ЭЭГ оказывают влияние также входы от лимбической системы -функциональной единицы, включающей гиппокамп, миндалину, мамиллярные тела и связанные с ними структуры. Лимбическая система вовлечена в организацию жизнедеятельности, эмоциональных реакций и мотивации. Нейрофизиологические исследования позволили также кое-что узнать относительно физиологической роли α-активности [1998]: α-ритм, по-видимому, модулирует и избирательно усиливает афферентные стимулы.
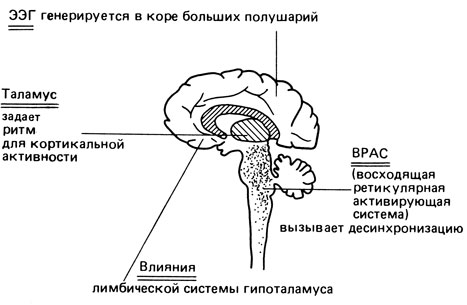
Рис. 8.24. Очень упрощенная схема человеческого мозга, показывающая структуры, участвующие в генерации ЭЭГ
Влияние наследственных вариаций ЭЭГ на личность. Личностные свойства и поступки индивида зависят от того, как справляется его мозг с информацией и насколько он спонтанно активен. Отсюда следует, что индивидуальные различия в нейрофизиологических параметрах такого рода должны приводить к психологическим различиям. Принимая во внимание нейрофизиологические данные, обрисованные в предыдущем параграфе, какое влияние на "личность" и умственную деятельность будут оказывать описанные выше варианты ЭЭГ? Можно предположить, что пробанды с мономорфными α-волнами являются "сильными модуляторами и усилителями", в то время как пробанды с низкоамплитудной ЭЭГ - слабыми. Считается общепринятым, что β-волны, особенно при их диффузном распределении (этот вариант, по-видимому, результат мультифакториального наследования) возникают вследствие высокого уровня тонической активации во ВРАС. Следовательно, у пробандов с этим вариантом ЭЭГ можно ожидать нарушения в модулирующей функции α-активности.
Результаты, полученные при обследовании 298 пробандов с различными вариантами ЭЭГ, вполне согласуются с этими предположениями [2231-2234].
1. Лица с мономорфными α-волнами в среднем проявляют себя активными, стабильными и надежными людьми. Пробанды с высокой вероятностью обнаруживают признаки высокой спонтанной активности и упорства; точность в работе, особенно в условиях стресса, и кратковременная память - самые сильные их качества. С другой стороны, переработка информации протекает у них не очень быстро.
2. Категория лиц с низкоамплитудной ЭЭГ демонстрирует низкую спонтанную активность; пробанды склонны быть экстравертами и ориентироваться на окружающих. Особенно хорошо развита у них пространственная ориентация. Недавно предполагавшиеся различия в переработке информации между группами 1 и 2 были прямо показаны при изучении усредненных вызванных потенциалов в ЭЭГ [2230].
3. В тестах, оценивающих концентрацию внимания и аккуратность, лица с диффузными β-волнами делают большое число ошибок, несмотря на низкую скорость работы. Устойчивость к стрессу представляется низкой. Нарушение α-механизма высокой тонической активацией ведет к нарушению выполнения тестов на интеллект, особенно тестов, оценивающих способность к пространственной ориентации.
Сильно упрощенная картина предполагаемых взаимоотношений между вариантами ЭЭГ и свойствами личности представлена в табл. 8.17. В ней перечислены и некоторые дополнительные варианты. Например, пробанды с быстрым вариантом затылочного α-ритма (16-19/с), по-видимому, превосходят других в абстрактном мышлении и в ловкости движений. Вероятно, они способны быстро перерабатывать информацию. В литературе по ЭЭГ есть сообщения о положительной корреляции между α-частотой и умственной деятельностью [2231].
![Таблица 8.17. Наследственные варианты ЭЭГ, их генетическая основа и психологические последствия [2231]](pic/000062.jpg)
Таблица 8.17. Наследственные варианты ЭЭГ, их генетическая основа и психологические последствия [2231]
Другой, очень редкий вариант ЭЭГ, не представленный в табл. 8.17, отличается от сходных на первый взгляд типов рядом свойств: например, волны частотой 4-5/с блокируются при открывании глаз и немедленно, после минутного перерыва, замещаются α-волнами. Генетическая основа этого варианта не совсем понятна; два случая конкордантных пар МЗ и небольшое число семей, имеющих более одного члена с такой ЭЭГ, указывают на генетические факторы, но большинство пробандов являются единственными носителями отклонения в своих, нормальных во всех других отношениях, семьях. Многие пробанды с этим вариантом ЭЭГ обнаруживают нарушения в сфере эмоций и в автономной нервной системе; среди психически больных людей этот вариант встречается намного чаще, чем в общей популяции [2138]. Все это можно объяснить аномалией функций лимбической системы.
Связь между α-волнами и пространственной ориентацией. В тестах, требующих зрительной ориентации в пространстве (разд. 8.2.2.2), средние показатели у женщин хуже, чем у мужчин. На рис. 8.19 показана одна стандартная задача, предназначенная для проверки пространственного восприятия. Было обнаружено, что эта способность особенно неразвита у больных с синдромом Тернера. Поскольку в зрительном восприятии участвует теменно-затылочная область коры, вполне возможна связь между α-активностью в затылочной области и выполнением заданий в тестах на пространственное восприятие. Некоторые исследования говорили о снижении пространственных способностей с уменьшением α- и увеличением β-активности, как это предполагалось на основании половых различий в этих характеристиках. Однако, четких доказательств пока нет.
Усредненные вызванные потенциалы в ЭЭГ. Мозг отвечает на стимул генерацией определенного вида волны. Реакция, однако, так слаба, что теряется в "шуме", производимом ЭЭГ покоя. Это препятствие можно преодолеть путем многократного повторения стимула (т. е. вспышки света или звука) и суммации реакций. При этом фоновый шум сглаживается и остается усредненный вызванный потенциал (УВП). Было обнаружено, что его характерная форма конкордантна у МЗ близнецов [2001], а некоторые свойства коррелируют с показателями интеллекта [2069], свойствами личности и подверженностью психическим болезням [2011]. Заметим, однако, что в использованных методиках и полученных результатах имеется много противоречий. Недавно в обстоятельной работе Фогеля (Vogel et al., в печати) корреляции свойств УВП с интеллектом подтвердить не удалось.
8.2.3.5. Генетика алкоголизма
Алкоголизм определяется как зависимость от алкоголя, которая приводит к потере трудоспособности. Превратится или не превратится пьющий человек в алкоголика, во многом зависит от окружающей среды. Если в обществе отсутствуют напитки, содержащие алкоголь, никто не сможет стать алкоголиком. Но даже в западных странах, где такие напитки имеются в изобилии, люди, которые становятся алкоголиками, часто страдают личностными расстройствами, не имеющими ни психологических, ни социальных объяснений. В склонности к алкоголизму существуют индивидуальные различия, которые могут быть детерминированы генетически.
Модели на животных. Влияние генетической изменчивости на восприимчивость к алкоголю продемонстрировано в экспериментах на мышах и крысах. В предпочтении алкоголя были обнаружены четкие различия между инбредными линиями [2182]. Показано, что эти различия связаны не только с особенностями метаболизма алкоголя, но и с количественными и качественными различиями в реакциях мозга на алкоголь. Эти результаты наводят на мысль о том, что генетические различия между людьми также следует искать на двух уровнях - в метаболизме алкоголя и в воздействиях на физиологию мозга.
Исследования с помощью классических методов: работы на близнецах и приемных детях. При изучении привычки к потреблению алкоголя у 174 близнецовых пар в Швеции (среди них 48 МЗ пар) между МЗ близнецами была обнаружена более высокая конкордантность, чем между ДЗ парами. Она распространялась и на характер потребления (систематическое или в виде "кутежей") и на количество принимаемого алкоголя [2097]. Сходные данные получены на МЗ близнецах, воспитывавшихся раздельно [2195].
Исследования с помощью классических методов: исследование семей, близнецов и приемных детей. Изучение семей пробандов, страдающих алкоголизмом, проводились довольно часто. Несколько лет назад Коттон [2024] проанализировал результаты 27 работ, опубликованных на английском языке, в которых представлены данные обследования семей 6251 алкоголика и 4803 неалкоголиков. Несмотря на то что не все эти работы равноценны, так как не всегда исследовался контроль, можно сделать некоторые общие заключения: почти у одной трети всех алкоголиков по крайней мере один из родителей страдал тем же пороком; в большинстве случаев (25%; в различных исследованиях значения колебались от 2,5 до 50%) алкоголиком был отец. По данным большинства авторов, родственники женщин-алкоголичек поражены в большей мере, чем родственники алкоголиков-мужчин. Это может свидетельствовать о том, что женщины чаще становятся алкоголиками по причинам внутреннего порядка, а у мужчин непосредственная причина алкоголизма нередко связана с окружающими обстоятельствами. Среди членов семей больных шизофренией и аффективными расстройствами алкоголизм встречался намного реже, чем в семьях алкоголиков. Отсюда следует, что повышенная склонность к алкоголизму не является общим признаком больных, страдающих основными психическими заболеваниями. Среди алкоголиков не удалось обнаружить какого-либо особенного типа личности.
Амарк [1997] исследовал в Швеции родственников 645 алкоголиков. Помимо алкоголизма он выявил также повышений риск "психопатии" и "психогенных психозов" и не обнаружил случаев умственной отсталости, эпилепсии или эндогенных психозов.
Разумеется, само по себе скопление алкоголиков в определенных семьях нельзя рассматривать как показатель генетической предрасположенности. Поэтому изучение семей было дополнено исследованием близнецов и приемных детей. Имеются три обстоятельные работы на близнецах: две выполнены в Швеции [2097; 2088] и одна - в Финляндии. Кроме более высокой конкордантности МЗ, чем ДЗ близнецов, данные этих исследований позволили сделать несколько более конкретные выводы. Например, по результатам, полученным в Финляндии, МЗ близнецы определенно чаще конкордантны по частоте злоупотреблений алкоголем и количеству выпитого, чем ДЗ близнецы; до тридцатилетнего возраста (но не позже) они более конкордантны по степени потери контроля над собой. При рассмотрении социальных последствий алкоголизма между МЗ близнецами не наблюдалось более тесного сходства, чем между ДЗ близнецами. Вопреки ожиданиям сравнение жизненных обстоятельств дискордантных МЗ пар не дало какой-либо информации о специфических факторах окружения как причине дискордантности.
Недостатки близнецового метода уже не раз обсуждались (разд. 3.8.4). Их можно избежать в исследованиях на приемных детях. Ряд таких исследований был выполнен в Дании [2229].
В табл. 8.18 сопоставлены результаты обследования 55 усыновленных датчан в возрасте от 20 до 40 лет, у которых по крайней мере один из биологических родителей был госпитализирован по поводу алкоголизма, и 78 контрольных лиц [2057; 2058]. Все обследованные входили в группу из 5483 приемных детей, исходно предназначенную для изучения шизофрении. Между приемными семьями пробандов и контрольных лиц различий в социоэкономических данных и в отношении к алкоголю не было. У сравниваемых групп не совпадала частота разводов, в то время как другие типы психопатологии были сходными. В отношении алкоголизма выборки различались только в том случае, если алкоголизм определять как расстройство, вызывающее трудности в социальных взаимодействиях и работе, связанное с потерей контроля, с абстинентным синдромом, включающим галлюцинации, и с психиатрическим лечением. Люди, просто много пьющие, встречались среди контрольных лиц примерно с такой же частотой. Это исследование было дополнено другим, в котором сравнили 20 сыновей алкоголиков, взятых на воспитание с 30 их родными братьями, которые росли в домах их собственных родителей [2057]. Как и ожидалось, социоэкономические условия в этих домах в среднем были хуже, чем в семьях приемных родителей. Результаты исследования, однако, оказались неожиданными: среди воспитывавшихся в собственных домах братьев доля алкоголиков была примерно такой же, что и в группе усыновленных. Если бы окружающая среда играла причинную роль в возникновении алкоголизма, можно было бы ожидать намного больше алкоголиков в группе неотданных на воспитание лиц. Эти данные говорят о том, что в обществе, в котором алкогольные напитки общедоступны, преимущественно генетические факторы определяют, станет данный индивид алкоголиком или нет. В аналогичном исследовании сравнивали дочерей алкоголиков, отданных на воспитание, с их сестрами, остававшимися с родителями-алкоголиками. Несмотря на выраженное различие в семейном окружении, алкоголизм в обеих группах встречался примерно одинаково часто и был более распространен, чем в популяции женщин в целом [2059].
![Таблица 8.18. Сопоставление проблем, связанных с пьянством, и характера пьянства в двух группах лиц, воспитывавшихся приемными родителями [2057; 2058]](pic/000063.jpg)
Таблица 8.18. Сопоставление проблем, связанных с пьянством, и характера пьянства в двух группах лиц, воспитывавшихся приемными родителями [2057; 2058]
* (Взрослые мужчины, воспитывавшиеся приемными родителями: по крайней мере один из биологических родителей госпитализировался по поводу алкоголизма.)
** (Р < 0,05.)
*** (Р < 0,02.
Генетическая изменчивость метаболизма алкоголя. Если генетическая изменчивость в предрасположенности к алкоголизму существует, то каков ее механизм? Ответ можно искать на двух уровнях: метаболизм алкоголя и действие алкоголя на мозг.
В исследованиях на близнецах было обнаружено существенное генетическое влияние на метаболизм алкоголя [2162]. На рис. 8.25 показаны два наиболее важных этапа в окислении этанола. Ключевыми ферментами в этих случаях являются алкогольдегидрогеназа (АДГ) и альдегиддегидрогеназа (АлДГ). Оба фермента синтезируются в печени. АДГ определяется тремя аутосомными локусами (ADH1, ADH2, ADH3). Гены ADH1 и ADH3 активны преимущественно в период внутриутробного развития; у взрослых за большую часть активности фермента в печени и почках отвечает ADH2. У 5-20% индивидов европейского происхождения и у 90% японцев был обнаружен атипичный вариант фермента. Поскольку при физиологическом значении рН он демонстрирует намного более высокую активность, чем обычный, было высказано предположение, что у его носителей окисление алкоголя происходит значительно быстрее. Кроме того, известно, что многие японцы "вспыхивают" после приема относительно небольшого количества алкоголя; лицо заливается краской, частота пульса возрастает, становится дурно. Аналогичный эффект можно вызвать у носителей более распространенного варианта алкогольдегидрогеназы, если вместе с алкоголем дать им дисульфирам (антабус). Известно, что это вещество увеличивает уровень ацетальдегида путем торможения активности фермента альдегиддегидрогеназы (АлДГ) (рис. 8.25), отсюда следует, что причиной прилива крови может быть повышение уровня ацетальдегида. АлДГ тоже обнаруживает в японской популяции генетический полиморфизм при частоте каждого из аллелей около 50%, в то время как у лиц европейского происхождения генетические варианты редки. Распространенный среди японцев полиморфизм связан с уменьшением активности АлДГ и, по-видимому, ответствен за часто наблюдающийся феномен "вспыхивания". Сочетание быстрого образования альдегида со сниженным его расщеплением служит, вероятно, причиной вспыхивания после приема алкоголя.
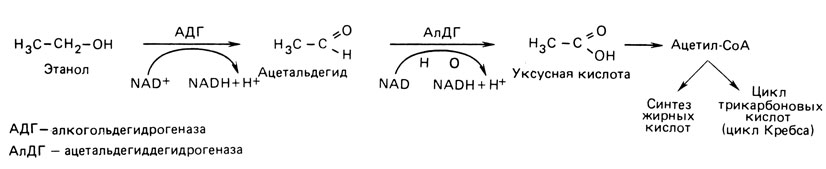
Рис. 8.25. Два последовательных этапа в деградации алкоголя. Первый этап находится под контролем алкогольдегидрогеназы, второй - под контролем ацетальдегиддегидрогеназы
Как отражается генетическое различие в метаболизме алкоголя на склонности к его употреблению? Как говорилось выше, прилив крови к лицу вызывает учащение пульса и определенный дискомфорт. Все это может удерживать носителя варианта АлДГ от слишком обильных возлияний и таким образом защитить от алкоголизма. Было показано, что среди японских алкоголиков этот вариант встречается намного реже, чем в других популяциях [2250].
Реакция мозга на алкоголь по показателям ЭЭГ. Гипотеза о том, что в основе алкоголизма лежат генетические факторы, появилась, когда реакции мозга на алкоголь стали исследовать с помощью ЭЭГ [2161; 1270]. Такая характеристика ЭЭГ, как амплитудно-частотные распределения в группах близнецов, показала высокую степень наследуемости: после приема алкоголя ЭЭГ МЗ близнецов становились еще более сходными. Еще важнее были различия в реакциях, обнаруживаемые при рассмотрении ЭЭГ покоя. Лица с выраженным и стабильным α-ритмом в состоянии покоя демонстрировали небольшие изменения после приема алкоголя (рис. 8.26). С другой стороны, лица, ЭЭГ которых в состоянии покоя характеризовалась менее развитыми α-волнами, обнаруживали более сильные реакции после приема алкоголя: их α-волны становились более выраженными и стабильными, чем в ЭЭГ покоя (рис. 8.27). Эти реакции у МЗ близнецов имели высоко конкордантный характер, но ДЗ близнецы по этому признаку иногда бывали дискордантны. Некоторые близнецы демонстрировали качественно иные реакции, например β-волн. Индивиды с низкоамплитудной ЭЭГ, доминантно наследуемым вариантом ЭЭГ (разд. 3.6.1), не реагировали на алкоголь увеличением генерации α-волн. Вероятно, существуют сильные, генетически определяемые различия между людьми в реакциях мозга на алкоголь. Эти различия не связаны с соответствующей изменчивостью метаболизма алкоголя. Корреляция между уровнем алкоголя в крови и реактивностью ЭЭГ отсутствовала. Еще более существенно то, что вызываемая алкоголем модификация структуры ЭЭГ сохраняется относительно долго и поддерживается после того, как метаболизм в основном завершен. Очевидно, генетически контролируемые различия характерны для реакции самого мозга.
![Рис. 8.26. Взрослые мужчины - члены МЗ близнецовой пары с хорошо развитым затылочным α-ритмом. Введение алкоголя в дозе 1,2 г/кг веса приводит к относительно небольшому увеличению α-активности через 120 мин [2161]](pic/000065.jpg)
Рис. 8.26. Взрослые мужчины - члены МЗ близнецовой пары с хорошо развитым затылочным α-ритмом. Введение алкоголя в дозе 1,2 г/кг веса приводит к относительно небольшому увеличению α-активности через 120 мин [2161]
![Рис. 8.27. Взрослые монозиготные близнецы мужского пола с относительно плохо выраженными а-волнами в ЭЭГ покоя; через 120 мин после приема 1,2 г/кг этанола α-ритм поразительно усилился [2161]](pic/000066.jpg)
Рис. 8.27. Взрослые монозиготные близнецы мужского пола с относительно плохо выраженными α-волнами в ЭЭГ покоя; через 120 мин после приема 1,2 г/кг этанола α-ритм поразительно усилился [2161]
Могут ли нейрофизиологические механизмы ЭЭГ объяснить различные реакции на алкоголь? Механизмы генерации ЭЭГ, которые были разобраны в разд. 8.2.3.4, помогают понять, как алкоголь изменяет работу мозга. У лиц с хорошо развитым α-ритмом его водитель ритма довольно слабо реагирует на воздействие. С другой стороны, у лиц, спонтанный α-ритм которых в норме склонен к десинхронизации, синхронизирующая функция водителя улучшается. Тенденция к десинхронизации может быть вызвана более сильным влиянием ретикулярной системы. Как связать эти факты?
Уже говорилось о том, что у некоторых алкоголиков α-ритм выражен плохо [2161]. Точнее, их α-ритм часто характеризовался параметрами, сходными с параметрами α-ритма близнецов, демонстрирующих сильные реакции на алкоголь (рис. 8.27). Не вполне ясно, была ли эта структура ЭЭГ причиной алкоголизма или следствием действия алкоголя на мозг - прямого или опосредованного повреждением печени. Известно, что приемы медитации типа трансцедентальной медитации облегчают α-активность. Субъективным следствием медитации является чувство расслабленности и умиротворенности. Такого же результата можно иногда добиться путем увеличения α-активности через биологическую обратную связь. Можно соединить ЭЭГ с устройством, которое будет издавать звук до тех пор, пока генерируются α-волны. Пробанду предлагают удерживать этот звук как можно дольше. Таким способом можно на ограниченное время усилить α-активность. Пробанды часто описывают свои ощущения при этом как расслабление, граничащее со счастьем; это состояние, по-видимому похоже на то, которое достигается при медитации [16].
Полученные результаты позволяют сформулировать предположение о нейрофизиологических и генетических основах некоторых случаев алкоголизма. Склонность к алкоголизму высока у лиц, обладающих в норме высоким уровнем тонической активации или слабой устойчивостью таламического водителя ритма. Уровень активации снижается под действием алкоголя, и человек чувствует себя лучше. Таким образом, потребление алкоголя сопровождается положительным подкреплением, что может приводить к алкоголизму. Это предположение было подтверждено в исследованиях ЭЭГ членов семей алкоголиков, которые не страдали алкоголизмом [2167; 2158]. Некоторые из них продемонстрировали сходную структуру ЭЭГ; отсюда следует, что ЭЭГ не могла быть результатом хронического злоупотребления алкоголем. В одной работе, однако, снижение среднего уровня α-активности было выявлено только у женщин и отсутствовало у мужчин, страдающих алкоголизмом. Согласно психиатрическим данным, алкоголизм этих женщин по большей части не связан с внешними обстоятельствами, в то время как большинство мужчин стало алкоголиками в результате социального давления и других внешних факторов [2167].
Этот пример показывает, как в зависимости от социокультурных условий окружающей среды генетически контролируемая восприимчивость может иметь разные фенотипические проявления. В западном обществе, где алкоголь считается обязательным фоном для общения, генетически восприимчивый индивид рискует стать алкоголиком. В буддистском окружении тот же человек, вероятно, целиком посвятил бы себя медитации.
Ослабление активации, вызываемой ретикулярной системой, не единственное следствие действия алкоголя на мозг. Исследования на близнецах говорят о том, что генетическая изменчивость может характеризовать и другие аспекты мозговой деятельности. Этот пример был описан столь подробно потому, что он представляет собой один из первых случаев, когда нейрофизиологические данные были использованы для развития генетической гипотезы. Нейрофизиология до сих пор не испытывала серьезного влияния со стороны генетики человека. В свою очередь ученые, занимающиеся проблемами генетики человека, очень редко предпринимали попытки включить в ход своих размышлений представления нейрофизиологии. Причина этого заключена в изолированности разных научных дисциплин. Основанием для особой изоляции физиологии может быть то, что она в большей мере, чем все другие биологические науки, имеет дело с объяснением работы интегрированных систем и механизмов обратной связи и редукционизм присущ ей в меньшей степени [267].
8.2.3.6. Физиология мозга: генетическая изменчивость нейромедиаторов
Анализ на биохимическом уровне: синапсы. Генетический анализ на уровне ЭЭГ вряд ли будет успешным, несмотря на то что концептуально он более удовлетворителен, чем анализ поведенческих фенотипов. Проблему необходимо решить на уровне ферментов и белков. Зададимся вопросом, генетическая изменчивость каких ферментов и белков может влиять на деятельность мозга?
Известно, что основными функциональными компонентами нервной системы являются нейроны [55]. Нейрон представляет собой клетку с одним ядром, одним длинным отростком, который называется нейритом или аксоном и служит эффекторным органом нейрона, и рядом сложно ветвящихся дендритов, образующих контакты с другими нервными клетками посредством так называемых синапсов. Рис. 8.28 демонстрирует основные органеллы синапса. Пресинаптическое окончание и постсинаптическая мембрана полностью изолированы друг от друга узкой синаптической щелью. Когда нервный импульс достигает пресинаптического окончания, передача через синапс осуществляется не электрическим, а химическим способом. Особые вещества, служащие передатчиками (медиаторами), упакованы порциями из нескольких тысяч молекул в пузырьках пресинаптических окончаний. Прибывающий импульс приводит к тому, что один или несколько пузырьков высвобождают молекулы передатчика (медиатора) в синаптическую щель. Таким образом, медиатор может подействовать на особые рецепторные участки постсинаптической мембраны. Это взаимодействие приводит к диффузии ионов Na+ через мембрану, что вызывает изменение электрического потенциала. Существуют два типа синапсов - возбуждающие и тормозные. Когда нейрон получает достаточное количество импульсов через возбуждающие синапсы, его аксон "срабатывает", т. е. генерирует импульс. С другой стороны, тормозные синапсы могут вызвать гиперполяризацию постсинаптической мембраны, которая не позволяет деполяризации достичь критического уровня, выше которого нейрон начинает генерировать импульсы. Таким способом возбуждающий импульс может передаваться другим возбуждающим нервным клеткам, число которых постоянно нарастает; "цепная реакция" не переходит во "взрыв", благодаря вставленным в цепочки тормозным нервным клеткам [55].

Рис. 8.28. Схематическое изображение адренергического синапса с его наиболее важными органеллами. Норадреналин синтезируется из тирозина, хранится в гранулах, высвобождается в синаптическую щель и связывается с рецептором эффекторной клетки на постсинаптической мембране (объяснения см. в тексте) (МАО - моноаминоксидаза; КОМТ - катехоламин-О-метилтрансфераза; ДБГ - дофамин-β-гидроксилаза
Именно в этой последовательности событий возможна генетическая изменчивость. Например, ферменты синтеза и расщепления молекул медиаторов могут обладать различной активностью, мембраны могут иметь структурные отличия, сказывающиеся на их проницаемости для молекул нейромедиаторов или ферментов, могут существовать различия в рецепторах и, наконец, на функции синапса могут оказывать влияние внешние регулирующие воздействия на разных уровнях. Самая простая возможность состоит в изменении количества молекул медиатора. В самом деле, некоторые результаты исследования психических заболеваний указывают на аномалии нейромедиаторной функции.
Химические типы нейромедиаторов (рис. 8.29). В качестве нейромедиаторов в мозге используется несколько соединений; синапсы специализируются на одном типе медиатора. Наиболее изученные на сегодняшний день медиаторы - норадреналин (адренергические синапсы) и ацетилхолин (холинергические синапсы). Этот факт можно объяснить чисто методическими причинами: указанные медиаторы можно исследовать в клетках периферической нервной системы. Например, нейроны симпатической нервной системы являются адренергическими, нейроны парасимпатической нервной системы - холинергическими. Однако в мозге эти два типа синапсов вместе принадлежат лишь небольшой части всех нейронов; в качестве нейромедиаторов здесь действует ряд аминокислот (гистамин, глутаминовая кислота, аспарагиновая кислота, глицин и другие). Существенным для синаптической активности является не только синтез, но и процесс инактивации медиатора. На рис. 8.29 представлены основные их типы.
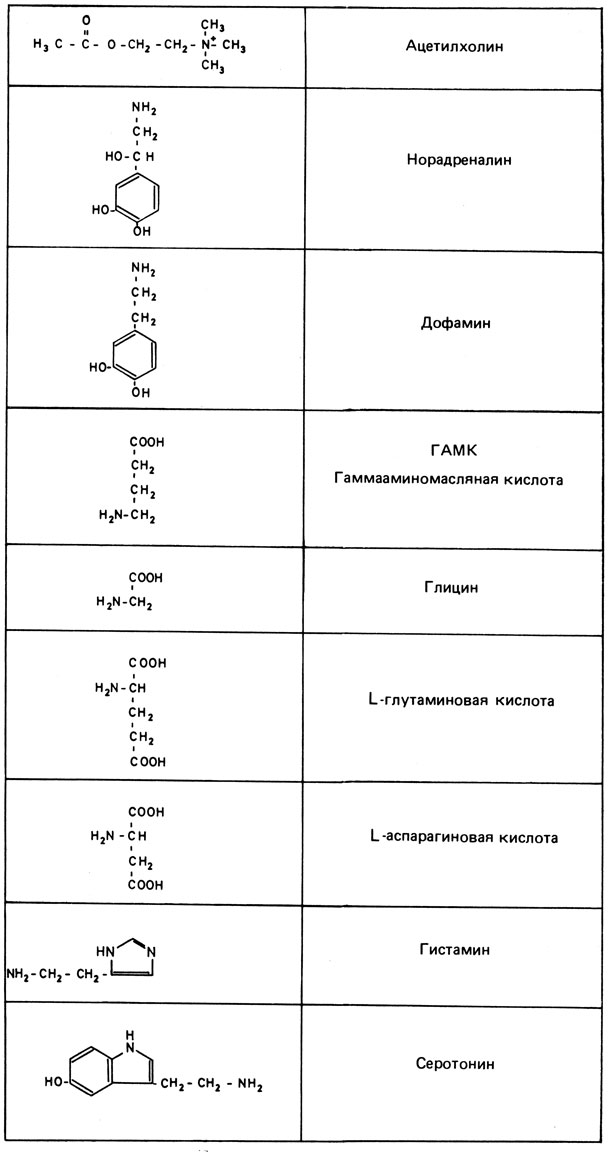
Рис. 8.29. Основные группы нейромедиаторов
Особый интерес вызывают два класса веществ, которые, по-видимому, изменяются при аффективных расстройствах и шизофренических психозах: катехоламины - норадреналин и адреналин с их предшественниками; индоламины - особенно 5-окситриптамин (серотонин). Мы ограничимся рассмотрением только одной группы - катехоламинов.
Катехоламины. Адреналин и норадреналин образуются из тирозина. Деятельность адренергического синапса представлена на рис. 8.28. Мы используем этот пример для демонстрации возможных мишеней генетической изменчивости и одновременно для демонстрации экспериментальных подходов к анализу этой изменчивости. В контексте нашей книги мы можем представить только очень упрощенную картину. Норадреналин, не использующийся как медиатор или уже выполнивший свою роль, должен быть инактивирован. Неоднократно изучалось участие двух ферментов в этом процессе - катехол-О-метилтрансферазы (КОМТ) и моноаминоксидазы (МАО). Путем изменения синтеза или деградации норадреналина его концентрация может быть повышена или снижена вплоть до полного исчезновения норадреналина в синапсах.
Генетический анализ изменчивости этих и других ферментов мозга трудно осуществить, так как человеческий мозг недоступен для прямого исследования. Есть два пути для преодоления этой трудности:
1) эксперименты на животных;
2) изучение аналогичных ферментов в других, более доступных тканях.
Эксперименты на животных по изучению генетической изменчивости метаболизма катехоламинов [2002; 2021; 2022]. Было обнаружено, что в надпочечниках мышей линии BALB/c активность ферментов тирозингидроксилазы, дофамин-Р-гидроксилазы и фенилэтаноламин-М-метилтрансферазы примерно вдвое выше, чем у другой инбредной линии - BALB/cN. При исследовании F1, F2 и потомства от возвратного скрещивания было выяснено, что активность этих ферментов контролируют единичные гены; это означает, что либо структурные гены этих ферментов тесно сцеплены, либо они находятся под общим регуляторным генетическим контролем.
Установлено, что превращение норадреналина в адреналин зависит от уровня стероидов, на него влияют гипофизэктомия или холодовый стресс. Период времени, за который соответствующий фермент осуществляет расщепление, неодинаков у разных линий. Кроме того, межлинейные различия у мышей могут существовать даже в механизмах контроля [2002].
Другим источником генетической изменчивости служит сАМР, который, как было обнаружено, действует в качестве вторичного посредника для различных гормонов и нейромедиаторов [120; 220]. Установлено, что содержание сАМР в мозге четырех инбредных линий мышей различается.
Эти эксперименты говорят о том, как сложна регуляция количества норадреналина в адренергических синапсах мозга; уместно напомнить, что связанные с этим различия в адренергической активности коррелируют с различиями в поведении (разд. 8.1.2). Учитывая всю сложность этих процессов, трудно представить себе, каким должен быть подход к исследованию ферментов мозга у человека и как на основании определения активности ферментов у животных делать выводы о возможных различиях нейромедиаторной функции в человеческом мозге. И все же есть надежда, что эксперименты на животных приведут нас к открытию генетической изменчивости и, следовательно, помогут разобраться в генетике нормального и отклоняющегося от нормы поведения.
Психотропные вещества [2169]. Психофармакологические препараты могут оказывать влияние на симптомы аффективных расстройств и психических заболеваний. Это обстоятельство стимулирует исследование механизмов психических болезней. Было обнаружено, что указанные вещества влияют на медиаторную функцию в синапсах, особенно на функцию норадреналина. Отмечалось, например, что одни больные депрессией лучше реагируют на ингибиторы моноаминоксидазы (МАО), а другие - на трйциклические антидепрессанты типа имипрамина. Более того, родственники пробанда, которые страдали депрессией, положительно реагировали на то же самое вещество, что и сам пробанд. Эта семейная тенденция отвечать на вещества преимущественно одного класса говорит о наличии генетической детерминированности. Оба соединения оказывают влияние на функцию норадреналина в адренергических синапсах; ингибиторы МАО ослабляют деградацию адреналина, увеличивая тем самым его количество в синапсах. Трициклические антидепрессанты, такие, как имипрамин, угнетают обратный захват адреналина выделившим его нейроном, увеличивая таким образом пригодное для нейропередачи количество адреналина. Семейные различия в терапевтической эффективности этих веществ могли бы указывать на разного рода генетические аномалии на уровне синапсов. Определенные заключения на этот счет сделать, однако, трудно, поскольку между людьми существуют генетические различия в метаболизме данных веществ и, следовательно, в уровне их содержания в крови. Такие различия особенно тщательно были исследованы для трициклического антидепрессанта нортриптиллина, который лишь немного отличается от имипрамина [2169]. При объяснении психофармакологических реакций с точки зрения генетических различий всегда нужно рассматривать оба аспекта - метаболизм вещества и мишень его действия - главным образом мозг. Крайне необходимы эксперименты, в которых уровень биосинтеза и концентрация в крови поддерживались бы постоянными с тем, чтобы можно было исследовать эффекты на уровне мозга. Исследования такого рода на людях необходимы не только для более глубокого понимания генетических основ аффективных и других психических заболеваний, но и для рационального их лечения с помощью фармакологических препаратов.
8.2.3.7. Аффективные расстройства и шизофрения
Генетическое изучение аффективных расстройств и шизофрении. Аффективные расстройства и шизофрения - это заболевания с ясно различимыми в большинстве случаев клиническими признаками. Генетические исследования этих состояний имеют длинную историю. После простого описания множества случаев заболеваний в предменделевский период в 1916 г. появилась классическая статья Рюдина [2186] (своей статистической основой обязанная сотрудничеству с Вайнбергом из знаменитой пары Харди - Вайнберг), которая стала образцом фенотипически-биометрического подхода к исследованию заболеваний такого рода. Следуя этому образцу, были проведены многие исследования на семьях и близнецах. С их помощью было четко показано, что генетическая предрасположенность занимает основное место среди причин аффективных расстройств и шизофрении (рис. 8.30 и 8.31). Кроме того, эта работа внесла существенный вклад в разработку более тонких статистических методов определения величин эмпирического риска, необходимых для генетических рекомендаций [744]. Однако многих исследователей эти эмпирические данные не удовлетворили, был предпринят целый ряд попыток, по большей части тщетных, продвинуть генетический анализ ближе к уровню действия генов, для того чтобы раскрыть биологические основы этих заболеваний.
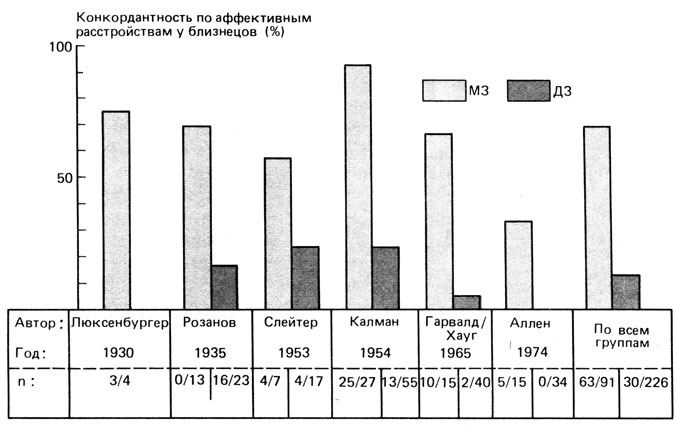
Рис. 8.30. Конкордантность и дискордантность по аффективным расстройствам у МЗ и ДЗ близнецов. Цифры 3/4 и т. д. обозначают соотношение общего числа близнецовых пар и числа конкордантных пар
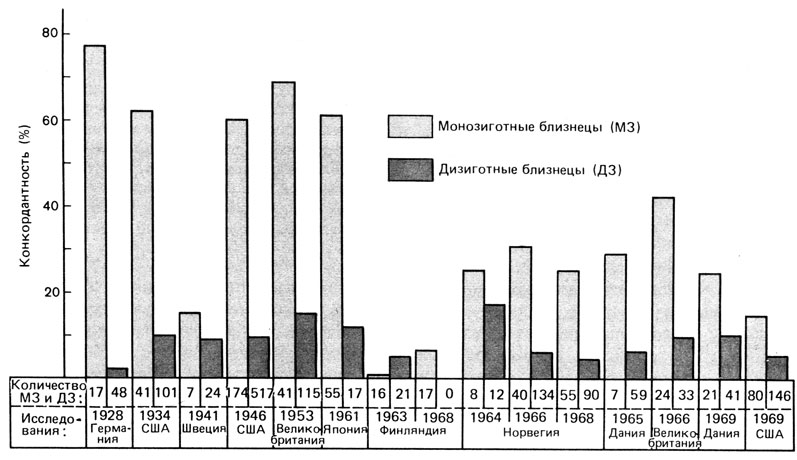
Рис. 8.31. Конкордантность и дискордантность по шизофрении у МЗ и ДЗ близнецов из разных стран
Аффективные расстройства включают маниакально-депрессивные или биполярные заболевания и униполярную депрессию. Другая большая группа широко распространенных психозов обычно классифицируется как шизофрения. Аффективные расстройства характеризуются главным образом циклическими изменениями настроения (депрессия или мания), в то время как при шизофрении основными симптомами являются отклонения в характере мышления и потеря связи с реальностью.
Близнецовые и семейные исследования при аффективных расстройствах [2252, 2051]. В более ранних исследованиях группу аффективных расстройств рассматривали как единое целое. Исследования на близнецах и семьях, направленные на определение величин эмпирического риска для родственников разной степени родства, начались с изучения случайно подобранных групп больных и близнецов (разд. 3.3.6). На рис. 8.30 представлены опубликованные данные по близнецам. Очевидно, что конкордантность между МЗ близнецами намного выше, чем между ДЗ близнецами, что указывает (при условии простой интерпретации данных) на вклад генетических факторов. Особенно важно то, что величина конкордантности для 12 МЗ пар, воспитывавшихся раздельно, составляла 67%, т. е. имела тот же порядок, что и конкордантность у МЗ близнецов, воспитывавшихся вместе [2252]. Но даже у МЗ близнецов конкордантность далека от полной, что свидетельствует о важности факторов окружающей среды. К сожалению, величины конкордантности на рис. 8.30 были подсчитаны без поправки на возраст. Поэтому не исключено, что некоторые из дискордантных пар со временем станут конкордантными.
Биполярный и униполярный типы: величины эмпирического риска. В близнецовых и семейных исследованиях больных с биполярными расстройствами, т. е. больных с чередующимися фазами мании и депрессии, не отделяли от больных, страдающих только депрессией (униполярные случаи). Леонгард [2114] впервые предположил, что с генетических позиций эти два расстройства могут отличаться. Последующие исследования подтвердили, что у биполярных пробандов родственники, страдающие биполярными расстройствами, встречаются чаще, чем у монополярных пробандов. Однако распространенность униполярной депрессии среди родственников биполярных больных намного выше, чем в общей популяции. Эти выводы были сделаны при сравнительном изучении биологических и приемных родителей биполярных больных [2131]. Установлено, что психопатология и особенно аффективные расстройства среди биологических родителей пробандов с биполярным психозом, воспитывавшихся приемными родителями, выражены в той же мере, что и среди родителей пробандов, воспитывавшихся в их родных семьях, тогда как частота психических расстройств у приемных родителей пробандов с маниакально-депрессивным психозом сходна с таковой у приемных родителей здоровых контрольных лиц.
Аналогичное исследование в случае депрессивных расстройств главным образом униполярного типа дало совершенно иной результат [2105]. Количество зарегистрированных психических заболеваний у приемных отцов было примерно в пять раз выше, чем в контроле (приемные отцы тщательно подобранных контрольных психически здоровых людей). С другой стороны, у биологических матерей пробандов женского пола было обнаружено только трехкратное увеличение частоты психических заболеваний. Эти данные указывают на важный вклад семейных экзогенных факторов в развитие униполярного аффективного заболевания. Кроме того, среди родственников больных аффективными расстройствами чаще наблюдались и некоторые другие психические нарушения, такие, как необычные колебания настроения, небольшая или умеренная депрессия, алкоголизм и острый нерецидивирующий психоз. Другой интересный аспект - половые различия. У биполярных пробандов близкие родственники женского пола в 1,5-2 раза чаще страдают психическими расстройствами, чем близкие родственники мужского пола. Среди близких родственников униполярных пробандов не было выявлено столь выраженных половых различий. В табл. 8.19 приведены наиболее важные величины эмпирического риска.
![Таблица 8.19. Величины эмпирического риска для аффективных расстройств (шесть групп) [2051]](pic/000071.jpg)
Таблица 8.19. Величины эмпирического риска для аффективных расстройств (шесть групп) [2051]
Недавно были предприняты попытки дальнейшего подразделения аффективных расстройств на биологически более однородные подгруппы. В замечательной работе Ангста [2000] выделены три подгруппы биполярного психоза в зависимости от того, преобладает ли депрессивная или маниакальная фаза или обе возникают примерно с одинаковой частотой. Самый высокий риск заболевания для близких родственников был обнаружен в последней группе. Кроме того, генетический риск возрастает с увеличением в семье пораженных заболеваниями родственников.
Простые типы наследования. На первый взгляд имеющиеся данные об эмпирическом риске несовместимы с простым типом наследования. Однако наблюдения довольно определенно говорят об участии, по крайней мере в некоторых случаях, главных генов. Описаны родословные, прямо указывающие на аутосомно-доминантный тип наследования. Есть такие, которые свидетельствуют о доминантном наследовании, сцепленном с Х-хромосомой; эти случаи характеризуются, по-видимому, ранним началом и хорошей реакцией на лечение литием [2245]. В таких семьях наблюдались биполярные и униполярные психозы как в тяжелой, так и в легкой, сомнительной форме. Отличить сцепленное с Х-хромосомой доминантное наследование от аутосомно-доминантного трудно (см. разд. 3.1.4).
Доводом в пользу локализации главного гена в Х-хромосоме могло бы стать обнаружение сцепления с Х-маркером, таким, как неспособность различать красный и зеленый цвета. Были приведены данные как в пользу сцепления с этим маркером, так и против него [2053]. Некоторое сомнение порождалось тем обстоятельством, что Xg-локус тоже обнаруживал сцепление, несмотря на то что он расположен далеко от локусов, определяющих неспособность различать красный и зеленый цвета (разд. 3.4.3). Относительно недавнее исследование продемонстрировало в некоторых родословных тесное сцепление с дефектами цветового зрения при отсутствии сцепления с Хg [2180]. Доля семейств с Х-сцепленным наследованием, по-видимому, невелика, так как в случайных сериях семей не было обнаружено уменьшения передачи от отца к сыну [2000].
Нет никаких данных о сцеплении с X-маркерами в семьях, где обнаруживаются только униполярные психозы. Кроме того, в этой группе женщины страдают психическими расстройствами намного чаще, чем мужчины. Болезнь у них начинается обычно в более позднем возрасте, депрессии возникают в периоды гормональной нестабильности, характерной для беременности, послеродового периода и особенно для менопаузы. Таким образом, данные семейных исследований аффективных расстройств, подтверждая предположение о значимом генетическом вкладе в развитие этих заболеваний, указывают на их генетическую гетерогенность и различные биологические механизмы.
Близнецовые и семейные исследования при шизофрении [173; 2061; 2060; 2252]. Результаты близнецовых исследований шизофрении суммированы на рис. 8.31. Демонстрируя в целом более высокую конкордантность между МЗ близнецами по сравнению с ДЗ, эти данные обнаруживают значительную степень статистической гетерогенности в конкордантности МЗ пар. Относительно недавние исследования выявляли, как правило, меньшую конкордантность, чем более давние работы. Это расхождение отчасти обусловлено способом оценки. Более ранние исследования основывались на сильно смещенных выборках (разд. 3.8.6). Пробандов выбирали из случайных групп больных; авторы выясняли, имеют ли эти пробанды близнецов и страдают ли эти близнецы психическими расстройствами. С другой стороны, исследования, выполненные недавно в скандинавских странах [2042; 2108; 2217], основаны на репрезентативных выборках из общей популяции. Сначала в популяции выявляли все близнецовые пары. Затем все пары, в которых по крайней мере один из близнецов страдал шизофренией, включали в исследование. Этот метод позволяет охватить пробандов с менее тяжелыми расстройствами, которые могут вообще не попасть в исследование, основанное на госпитальных выборках. Низкие величины конкордантности в таких работах не удивительны. О низких величинах конкордантности для МЗ близнецов с мягкой клинической симптоматологией неоднократно сообщали и другие авторы [2236].
Анализ дискордантных МЗ пар выявил межличностные различия в преморбидном периоде - в детстве и юности [2217, 2218]. В табл. 8.20 представлены некоторые факторы окружающей среды, исследованные в работе на близнецах, проведенной в Финляндии. Описание четырех идентичных близнецов Дженьян (разд. 8.2.1.5) иллюстрирует, как, несмотря на идентичную генетическую конституцию, различия в состоянии здоровья близнецов, в отношении к ним родителей и сибсов, в сочетании с внешним стрессом приводят к тяжелым психозам одних, тогда как другие либо остаются здоровыми, либо страдают менее сильным расстройством. Один из аргументов против генетической интерпретации конкордантности между МЗ близнецами, выросшими вместе, основан на их тесном социальном взаимодействии, которое могло бы усиливать шизофреническую симптоматологию у второго близнеца при наличии заболевания у первого (разд. 3.8.4). Однако существуют данные по меньшей мере по 12 близнецовым парам, воспитывавшимся раздельно, в которых по крайней мере один из близнецов заболел шизофренией; в 7 парах другой близнец тоже был шизофреником [2061]. Эта цифра хорошо соответствует величинам, приводимым в исследованиях МЗ близнецов, воспитывавшихся вместе (рис. 8.31). Другим аргументом могло бы быть то, что родители, страдающие шизофренией, создают особенно плохие условия для умственного развития ребенка и тем самым увеличивают для него риск тоже стать шизофреником. Ответ на этот вопрос можно получить при сравнительном исследовании детей дискордантных МЗ. Если присутствие или отсутствие родителя, страдающего шизофренией, является существенным фактором, то детей клинически здорового близнеца болезнь будет поражать реже, чем детей близнеца с шизофренией. Исследование, проведенное в Дании, не обнаружило такой закономерности и, следовательно, подтвердило справедливость генетической интерпретации [2040]. Анализ характера взаимодействий между родителями и близнецами показал, что в отношении МЗ близнецов их родители ведут себя, как правило, не более сходно, чем родители ДЗ близнецов, по отношению к своим детям. Это касается спонтанного поведения, но их реакции на МЗ близнецов похожи в большей мере, потому что эти близнецы ведут себя в большей степени одинаково [2102].
![Таблица 8.20. Факторы среды для 16 МЗ близнецов, дискордантных по шизофрении [2217]](pic/000072.jpg)
Таблица 8.20. Факторы среды для 16 МЗ близнецов, дискордантных по шизофрении [2217]
Величины эмпирического риска. Величины эмпирического риска для шизофрении предполагают следующие заключения (см. также табл. 3.7, разд. 3.3.6).
1. У близких родственников шизофреников риск развития болезни в 10-20 раз выше, чем в общей популяции. Абсолютная величина риска составляет около 10-15%.
2. У родственников пробандов с кататонической или гебефренической симптоматологией риск выше, чем у родственников пробандов с параноидной, а также простой шизофренией.
3. Внутри семей существует корреляция клинических форм заболевания. Родственники больных с кататонической формой имеют больший риск заболеть кататонической шизофренией, чем родственники больных параноидной шизофренией. Однако у родственников больных кататонической и гебефренической формами риск заболеть параноидной шизофренией также увеличен по сравнению со средней величиной в популяции.
4. У сибсов пробандов риск минимален (около 10%), если оба родителя пробанда здоровы; он повышается до 15%, когда болен один из родителей, и становится еще выше (40-60%), когда больны оба.
5. Среди родственников шизофреников часто наблюдается тип личности, хотя и не явно патологический, но отклоняющийся от нормы и классифицируемый как шизоид [19].
Характерный пример величин риска представлен в табл. 3.17, разд. 3.3.6. Накопление в семьях психических расстройств не обязательно означает участие генетических факторов. К сходному результату могла бы привести патогенная семейная обстановка. Психиатры активно ищут такие семейные факторы. Результаты исследований на близнецах дают более веские свидетельства в пользу генетически контролируемой подверженности. Однако полезность данных по близнецам, особенно в области генетики поведения, по нашему мнению, можно оспаривать (см. разд. 3.8.4). Наилучший способ разделения вклада генетических и средовых влияний состоит в исследовании приемных детей в сопоставлении их с биологическими и приемными родителями.
Исследование шизофрении у лиц, воспитывавшихся приемными родителями. Первое исследование такого рода по шизофрении было опубликовано Хестоном в 1966 году [2072]. Он исследовал взрослых, которые в детстве были отданы на воспитание и биологические матери которых страдали шизофренией. Пробанды были отделены от больных матерей, не имели с ними никаких контактов и не жили с родственниками по материнской линии.
Пятеро из 47 исследованных потомков были поражены шизофренией. Такая же частота наблюдается у детей, живших со своими родителями, которые страдали шизофренией. С другой стороны, ни один из 50 индивидов, составлявших соответствующую контрольную группу усыновленных лиц с родителями, не болевшими шизофренией, не стал шизофреником. Та половина детей (от матерей с шизофренией), которые не были больны в явной форме, обнаружили высокую психосоциальную неполноценность; другую половину составляли весьма преуспевающие взрослые люди.
Более тщательные исследования были выполнены коллективом американских и датских ученых в Дании. В этой стране хорошо поставлен учет, и можно получить надежные данные по усыновлению и шизофрении. Было обнаружено, что частота шизофрении среди биологических родственников приемных пробандов-шизофреников примерно в три раза превышает частоту заболевания среди их приемных родственников, тогда как у приемных родителей пробандов шизофрения наблюдалась не чаще, чем у приемных родителей здоровых приемышей (они чаще демонстрировали другие типы психологических отклонений). Кроме того, риск заболеть шизофренией для детей, биологические родители которых страдают шизофренией, оказался одинаковым независимо от того, воспитывались ли они своими родными или были усыновлены в раннем детстве. Внутриутробные и перинатальные влияния от биологических матерей больных были исключены путем исследования усыновленных, которые были полусибсами шизофреников по отцовской линии и, следовательно, имели общую наследственность только с отцом. В этой группе шизофрения также наблюдалась чаще, чем в контрольной (приемыши, не имеющие больных шизофренией полусибсов или других родственников) [2104; 2184; 2241; 2242].
Биологические гипотезы шизофрении. Несмотря на все усилия, которые предпринимались исследователями в течение многих десятилетий, успехи в биологическом изучении шизофрении невелики. Анализировать данные трудно, потому что отсутствуют качественные или количественные критерии, по которым можно диагностировать шизофрению; более того, диагностические стандарты часто различаются у разных психиатров и в разных странах.
Все данные, полученные в исследованиях шизофрении на близнецах, в семьях и у приемных лиц, самым простым способом можно объяснить с помощью модели мультифакториального наследования в сочетании с пороговым эффектом. Эта формулировка не исключает возможности того, что в некоторых семьях в генетическую предрасположенность вносят свой вклад главные гены, так как вероятна генетическая гетерогенность между семьями. На предварительном уровне генетического анализа определяющую роль играет гипотеза о предрасположенности к стрессу [19; 2060]: стрессовые ситуации, которые преодолеваются большинством индивидов или приводят у них к "невротическим" симптомам, у людей, предрасположенных генетически, могут вызывать шизофренический психоз. Это объяснение, однако, не совсем удовлетворительно. Вопрос состоит в том, какие генетически определяемые физиологические изменения увеличивают риск заболевания шизофренией? Как и в случае аффективных расстройств, имеющиеся в настоящее время гипотезы сконцентрированы на аномалиях в метаболизме нейромедиаторов [2200]. Например, метионин может вызывать у хронических шизофреников острые психотические реакции. Известно, что психотомиметическое вещество мескалин является метилированным производным дофамина - предшественника норадреналина. Возможно, что у некоторых больных шизофренией патологически повышено метилирование дофамина и метионин усиливает эту патологию, являясь источником метильных групп. Другими возможными кандидатами на роль эндогенных психотоксинов являются метилированные производные серотонина (5-окситриптамин). Метаболизм индолов в целом и серотонина, в частности, находится в настоящее время в центре внимания многих работ в этой области. Например, может быть снижено поглощение мозгом предшественника серотонина - триптофана; такая гипотеза согласуется с эффектом метионина, поскольку метионин конкурентно блокирует поглощение триптофана. Другая возможность заключается в нарушении баланса между дофаминовой системой, активность которой повышена, и серотониновой системой, активность которой снижена. Это нарушение аналогично тому, которое наблюдается при болезни Паркинсона, когда повышенная активность ацетилхолиновой системы сочетается со сниженной активностью дофаминовой системы (см. перечень нейромедиаторов на рис. 8.29). Комбинированное введение триптофана с ингибиторами МАО (для предотвращения деградации серотонина) действительно приводит к улучшению состояния у некоторых больных шизофренией.
Эти примеры показывают, в каком направлении развивается мысль и идут эксперименты. Однако на сегодняшний день мы не располагаем какими-либо убедительными данными относительно биологических механизмов шизофрении. В настоящее время обсуждаются и другие гипотезы, например, затрагивающие свойства мембран и аномальные иммунные процессы. Результаты исследования близнецов и семей убедительно свидетельствуют о мультифакториальной генетической модели и генетической гетерогенности. Следовательно, во многих случаях подталкивать индивида к порогу психотического состояния и к психозу может не одна крупная биохимическая аномалия, а сумма нескольких или даже сумма нескольких небольших отклонений в сочетании с внешним стрессом. В других случаях решающей может быть одна крупная аномалия. История соматических теорий шизофрении не внушает оптимизма [2103]. Напрашивается простой вопрос: "Что такое шизофрения? Действительно ли она существует как единое заболевание?"
"Шизофрения" в свете генетики человека. Известно, что диагностика шизофрении крайне трудна и. часто диагноз основан на довольно спорных критериях. В разд. 3.8.14 обсуждался вклад генетики человека в теорию болезней в более общем плане. Речь шла о том, что представление о единой основной причине заболевания возникло в последние десятилетия девятнадцатого столетия, когда ведущей областью медицинской науки была бактериология. Эта концепция продемонстрировала свою силу, когда удалось открыть микроорганизмы, вызывающие туберкулез и сифилис. Без представления о главной причине болезни успех специфической терапии с помощью химических препаратов или антибиотиков был бы невозможен.
Диагноз раннего слабоумия по Крепелину (названного впоследствии Блейлером шизофренией) в свете такой концептуальной модели основывался на комбинации ряда клинических симптомов с постепенным ухудшением состояния больного, выявляемым при длительном наблюдении. Это диагностическое понятие безоговорочно подразумевало одну главную общую причину. Генетики интуитивно симпатизируют такой идее, потому что их идеальное представление о болезнях базируется на "врожденных ошибках метаболизма" Гэррода [75] или, еще конкретнее, на гемоглобинопатиях, где одна специфическая мутация определяет одно отклонение в белке, которое ведет к одной характерной болезни (разд. 4.3.2). Мыслящие психиатры, такие, как Джаспер или Блейлер, понимали, что слишком прямолинейное применение такой модели к этой группе заболеваний может привести к ложным заключениям. Однако крепелиновское представление о болезни оказалось удивительно живучим. Его не поколебали открытия последующих десятилетий. Выяснилось, что у многих больных не наблюдается ухудшения, а значительная часть случаев ухудшения (но не все) представляет собой артефакт, вызванный долговременной госпитализацией [19]. Сохранение этого диагностического понятия было обеспечено, по крайней мере отчасти, интересной стратегией: всякий раз, когда характерные для шизофрении симптомы наблюдались вместе с признаками органического заболевания, от диагноза шизофрении воздерживались и вместо него ставили такой диагноз, как "шизофреническая реакция". Когда все больные с шизофреническими симптомами, проявлявшие вдобавок признаки определенного органического заболевания, были исключены, осталась группа болезней, для которых не смогли обнаружить специфических причинных факторов.
В значительной части случаев органического поражения мозга шизофреноподобные симптомы описывались чаще, чем можно было ожидать на основе простой вероятности [2164]. Кроме того, существует много сообщений об атрофии мозга у хронических шизофреников. Правда, во многих из этих случаев при обследовании семей не обнаружили генетической предрасположенности к "истинной" шизофрении и настоящей причиной этого состояния было развитие иной болезни. С другой стороны, трудно не сделать вывода, что "... если бы диагноз органического поражения не был поставлен независимо от психиатрической симптоматологии, большинство случаев, несомненно, было бы отнесено к шизофрении" [2102].
Существует также ряд вполне определенных генетических состояний, которые сопровождаются шизофреноподобными психозами (табл. 8.21). К ним относятся 45,ХО- и XYY-кариотипы, различные липидозы взрослых, врожденная гиперплазия надпочечников, гомоцистинурия, болезнь Вильсона и некоторые другие [2164]. Некоторые случаи позволяют даже предположить вероятные биологические механизмы; например, снижение поступления фолиевой кислоты или изменение метаболизма и функции сульфатированных аминокислот типа метионина. Такие больные имеют, возможно, аномально высокую способность метилирования дофамина, и метионин, являясь общим источником метильных групп, может усиливать это действие.
Большинство шизофреников нельзя отнести к разряду больных, имеющих ясно очерченное органическое поражение генетической или негенетической природы. Однако во многих случаях были описаны легкие функциональные отклонения, такие, как уменьшение активности МАО и других ферментов, "рваный" ритм в сочетании с некоторым уменьшением α-активности в ЭЭГ или небольшими отклонениями в ЭЭГ-потенциалах, вызванных зрительными или слуховыми стимулами. Однако ни одно из этих отклонений нельзя найти у всех шизофреников, и потому их причинная связь с заболеванием всегда остается под вопросом. Дополнительное осложнение вносит тот факт, что большинство больных, исследуемых впервые, лечились психотропными препаратами, которые могли изменить многие из представляющих интерес параметров.
Возникает и другой вопрос: некоторые из состояний, перечисленных в табл. 8.21, характеризуются аутосомно-рецессивным типом наследования; в доминантных заболеваниях типа порфирии клинические симптомы проявляются только при особых стрессовых условиях. Не может ли быть так, что гетерозиготы (по метахроматической лейкодистрофии) или гомоцистинурии в большей мере подвержены заболеванию шизофренией, в особенности если эта генетическая "слабость" сочетается со склонностью к другим заболеваниям или если к ней добавляются стрессирующие факторы соматической или психологической природы? Возможное небольшое увеличение риска в отношении психозов у гетерозигот по фенилкетонурии отмечалось в разд. 8.2.1.3. Результаты семейных и близнецовых исследований шизофрении согласуются с мультифакториальной моделью наследования (разд. 3.6), включающей стрессорные факторы окружающей среды. Такая генетическая модель пригодна только для описания данных на предварительном уровне: она скорее ставит вопросы, чем отвечает на них. В случае шизофрении ответы будут проще, если представление о единой причине заболевания, введенное Крепелином и его последователями, заменить представлением о многопричинности этой болезни. Человеческий мозг является сложной системой, в которой взаимодействует великое множество структурных и биохимических подсистем. Его реакция на экзогенные и эндогенные стрессорные факторы зависит от индивидуальных, генетически контролируемых отклонений в пределах этих подсистем, от анамнеза жизни индивида, от вида и локализации стресса. Многие комбинации внешних и внутренних стрессорных факторов могут привести к одному и тому же или к похожему конечному результату: по-видимому, мозг обладает лишь ограниченным набором способов реагирования на такой стресс, а реакция зависит от специфических подсистем, на которые направлен стресс, и от их врожденной чувствительности.
![Таблица 8.21. Генетические расстройства, сопряженные с повышенным риском развития шизофреноподобных психозов (по [2164] с сокращениями)](pic/000073.jpg)
Таблица 8.21. Генетические расстройства, сопряженные с повышенным риском развития шизофреноподобных психозов (по [2164] с сокращениями)
От вопроса о причинах шизофрении следует перейти к вопросу о ее патогенезе. Какой из функциональных механизмов мозга изменяется при этом заболевании и каким образом изменения вызывают клинические проявления и симптомы? До сих пор ни одна из более или менее стройных теорий не завоевала симпатий большинства специалистов. Однако многие полагают, что общий конечный путь, в котором объединяются все этиологические факторы, мог бы иметь что-то общее с дисфункцией внимания или "ошибочной фильтрацией информации" [2124]. Стоит только возникнуть отклонению от нормы, включается в дело врожденная склонность к сохранению приобретенного. Эволюция снабдила человеческий мозг способностью "учиться" (разд. 7.2.4). Это означает, что характер функции изменяет структуру связей между нейронами таким образом, что повторение этой функции будет проходить легче. В нормальных условиях эта способность приносит определенную пользу, она помогает индивиду справиться с требованиями множества разнообразных обстоятельств окружающей среды. Но, подобно многим другим биологическим приспособлениям, она может стать гибельной при особых условиях, т. е. когда функция, предложенная для "изучения", имеет разрушительный характер.
Сходный механизм достаточно убедительно продемонстрирован для другой болезни мозга (или группы болезней) - эпилепсии. Каждый судорожный приступ в этом случае можно рассматривать как вспомогательный процесс при подготовке следующего. По-видимому, эпилептический приступ - еще один из очень немногих способов нашего мозга отвечать на различные стимулы. Большую часть принципов, достаточно подробно обсуждавшихся в этом разделе по шизофрении, можно было бы сформулировать сходным образом и при использовании в качестве примера этиологии и патогенеза эпилепсии [1999].
Стратегия исследований, направленных на выяснение генетических основ шизофрении. Как говорилось выше, понятие о шизофрении как заболевании является феноменологическим. В отличие от туберкулеза или фенилкетонурии оно не подразумевает единственной главной причины, что вовсе не умаляет значение этого понятия для диагностики. Анализ накопления в семьях, а также полезность этого понятия при генетическом консультировании способствуют лучшему пониманию причин заболевания. Для дальнейшего изучения генетики шизофрении целесообразна следующая стратегия.
1. Исследование "мультиплексных семей" (семей с несколькими пораженными болезнью членами). Такие исследования предоставляют наилучшую возможность открыть главные гены и, следовательно, главные биохимические аномалии.
2. Долговременные исследования детей группы высокого риска (т. е. детей, имеющих родителей шизофреников) обещают дать ценную информацию. Они уже осуществляются рядом авторов [2032; 2049; 2084; 2130].
3. Следует изучать нормальную генетическую изменчивость тех параметров, которые предположительно могут иметь важное значение для шизофрении.
4. В некоторых семьях неблагоприятная комбинация небольших количественных отклонений в биохимических и структурных показателях может вызывать заболевание приблизительно тем же способом, как было описано для косоглазия (разд. 3.6.2.7).
Ферменты нейромедиаторов и генетическая изменчивость нормального поведения. Нейромедиаторные ферменты обнаруживают отклонения в активности не только при аффективных расстройствах и психозах; изменчивость в заметных пределах существует также между нормальными индивидами. Близнецовые исследования показали, что эта изменчивость в значительной мере определяется генетически [2246; 2029]. Для одного из ферментов, дофамин-β-гидроксилазы, на основе семейных данных удалось продемонстрировать единый генетический вариант с нулевой активностью в сыворотке [2238; 2240]. Однако до сих пор не было сообщений о его корреляциях с поведенческими показателями или с функцией автономной нервной системы (синапсы симпатической нервной системы являются адренергическими).
Нейромедиаторы, подобно многим другим биологически активным молекулам, должны прореагировать с рецепторами (обычно на поверхности клеток), для того чтобы выполнить свою биологическую функцию. Эти рецепторы, как и ферменты, могут характеризоваться генетической изменчивостью. Большинство из известных до сих пор заболеваний, связанных с рецепторами, возникают из-за снижения рецепторной активности почти до нуля (разд. 4.6.4), но с помощью близнецовых исследований для адренергических рецепторов тромбоцитов человека [2165] и для рецепторов липопротеинов низкой плотности была установлена генетическая изменчивость в нормальном диапазоне. Рецепторы так же, как ферменты, можно изучать с помощью различных физико-химических методов. Вполне возможно, что именно такой путь исследований откроет новый, функционально важный тип генетической изменчивости.
Другие параметры биохимии мозга, которые могли бы влиять на его функции и поведение человека. Мозг обладает очень высокой потребностью в энергии. Он использует около 20% всего поглощаемого кислорода. Кроме того, мозг чрезвычайно чувствителен к отсутствию кислорода: достаточно нескольких минут для того, чтобы вызвать его необратимое повреждение. В связи с этим было бы разумно ожидать наличия генетической изменчивости у гликолитических ферментов ткани мозга человека [2149]. Однако полиморфизма у гликолитических ферментов обнаружено не было.
Критическая оценка попыток связать поведенческую изменчивость с биохимическими различиями в функции мозга. Несмотря на интересные предположения, попытки объяснить нормальное или патологическое поведение с помощью какого-либо биохимического механизма, детерминируемого генетически, пока не удались. Нет ни единой модели, на которой можно было бы выяснить цепь событий от генетического варианта до поведенческого признака. И все-таки этот подход заслуживает внимания. Учитывая нынешнее состояние исследований, можно предположить, что прорыв произойдет в области изучения аффективных расстройств.
Напомним, что норадреналин, дофамин и серотонин составляют только часть всех нейромедиаторов; для других соединений этого класса генетических исследований выполнено мало или они вообще не проводились. Нейромедиаторы представляют только один из аспектов мозговой функции, между тем индивидуальные различия вполне возможны и в развитии мозга, и в количестве нервных клеток, и в числе связей между ними. Например, число апикальных дендритов в зрительной зоне коры мозга мышей уменьшается, если животное растет в темноте [55; 2225]. Затылочный α-ритм в ЭЭГ в среднем хуже развит у взрослых людей, родившихся слепыми [16]. Как отмечалось выше, в нормальных условиях межиндивидуальная изменчивость ЭЭГ и особенно развитие α-ритма определяются исключительно генетическими факторами [921]. Однако структуры мозга, продуцирующие ЭЭГ зрительной коры, могут адекватно развиваться, только если они получают соответствующий сенсорный вход. Генетическая программа развития мозга может быть выполнена только при взаимодействии с окружающей средой. Сравнивая наши попытки проникновения в суть генетической изменчивости функции мозга, влияющей на поведение человека, с нашими знаниями о генетическом аппарате эритроцитов (разд. 4.2, 4.3), мы ясно понимаем, насколько фрагментарна информация относительно поведения. Если для гена гемоглобина мы можем шаг за шагом проследить, как определенное изменение в последовательности ДНК трансформируется в изменение фенотипа, в области генетики поведения у нас в руках только измерения и сравнение фенотипических показателей, которые далеки от действия генов. Мы делаем всего лишь первые шаги в анализе промежуточных переменных. Не окажется ли так, что участвующие здесь механизмы более сложны и запутанны, а взаимодействие со средой более существенно для адекватного развития функции мозга и даже его структурных элементов? В этом не было бы ничего удивительного, поскольку человеческий мозг является самым поздним и самым сложным продуктом эволюции. Немногие имеющиеся на сегодня данные поддерживают такое предположение [55].
В начале этой главы мы отметили, что генетика поведения представляет собой концептуально самое важное направление генетики человека. Однако наши знания в этом вопросе все еще в высшей степени неудовлетворительны и фрагментарны, теоретические позиции наименее продуманы, а у научных гипотез мало объективных оснований. Ученые - это люди со своими предубеждениями и эмоциями; когда нет разработанной теории и прочного эмпирического фундамента, они в большей мере подвержены влиянию своих личных симпатий. Наиболее ожесточенные споры породила проблема существования генетически детерминируемых различий поведения различных этнических групп.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'