IV.1. Независимое наследование взаимодействующих генов
IV.1.1. Расщепления при аутосомной локализации генов
Из работы Менделя следовало, что расщепления в F2 по нескольким генам, локализованным в аутосомах, подчиняются формуле независимого наследования (3:1)n, где n - число генов, по которым идет расщепление. Для двух генов это - 9АВ:3Ab:3аВ:1ab, для трех - 27АВС:9АВс:9аВС:9АbС:3Аbс:3аВс:3аbС:1abc. Из курса общей генетики известны примеры расщеплений для двух (9:3:3:1, 9:6:1, 9:3:4, 9:7, 12:3:1, 13:3, 15:1) и трех (27:37, 63:1 и др.) взаимодействующих генов.
Расщепления по нескольким генам часто трудно понять "на глаз". Для их расшифровки и объяснения полезно напомнить следующее. Количество классов в F2 может служить отправной точкой для выявления числа генов, по которым оно происходит: при моногенном различии между формами наблюдается либо два, либо три класса в F2. Появление в потомстве F2 четырех или большего числа фенотипических классов указывает на участие в расщеплении более одной аллельной пары (при гомозиготности исходных родительских форм). Соотношение классов, отличное от моногенных (3:1, 1:2:1), также свидетельствует об этом.
Для выявления соотношения фенотипов, полученного в опыте, можно предложить простой прием, пригодный для случаев предполагаемого независимого наследования. Он состоит в следующем. Последовательно проверяют гипотезы об участии в расщеплении двух, трех и т. д. генов. В соответствии с предложенной гипотезой о числе генов выборку, полученную в опыте, делят на число возможных сочетаний гамет, определяемое по формуле 4n, и таким образом находят величину одного сочетания. Затем оценивают расщепление в опыте, для чего делят число потомков в каждом классе на эту величину. Если полученное при этом соотношение классов в расщеплении похоже на ожидаемое в соответствии с предложенной гипотезой, то проводится обычная статистическая оценка отклонения по методу χ2. Если соотношение, полученное таким способом, невозможно объяснить действием предполагаемого числа генов или проверка по χ2 отвергает нулевую гипотезу, выдвигается новая гипотеза, например, об участии в расщеплении большего числа генов или об их сцепленном наследовании, которая также проверяется статистически.
В задаче № IV. 1 показана "рецептура" такого анализа.
В реципрокных скрещиваниях коричневых и сапфировых гомозиготных норок в F1 все щенки оказались коричневыми, а в F2 произошло расщепление: 46 коричневых, 5 сапфировых, 12 алеутских и 11 платиновых. Объяснить расщепление, определить генотипы исходных форм и гибридов F1.
Поскольку в F1 реципрокных скрещиваний наблюдается единообразие, признак не сцеплен с полом и исходные формы, по-видимому, гомозиготны. В F2 возникло четыре фенотипических класса. Это дает основание предположить, что родительские формы различаются более чем по одной аллельной паре, например, по двум генам. Величина одного сочетания гамет равна - 74:16 = 4,6. Определим примерное расщепление в опыте - 46:4,6 = 10 (~9), 12:4,6 = 2,6 (~3), 11:4,6 = 2,4 (~3) 5:4,6 = 1,1 (~1). Проверяем соответствие гипотезе о расщеплении по двум независимо наследуемым, комплементарным генам; χ2 = 1,4, p>0,60. Отклонение случайно, гипотеза не отвергается.
Исходные формы различаются по двум комплементарно взаимодействующим, независимо наследуемым генам. АВ - коричневые, Аb (или аВ) - алеутские, аВ (или Аb) - платиновые и ab - сапфировые; генотип исходных форм 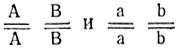 гибридов
гибридов 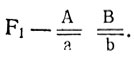
Очевидно, что четыре класса в расщеплении по одному признаку могут возникнуть и при взаимодействии трех и более генов. Однако характер расщеплений при этом должен быть иным. Например, при участии в расщеплении трех взаимодействующих генов возможны соотношения фенотипов: 27:27:9:1 - 27АВС:27(9АВс + 9АbС + 9аВС):9(3Abc + 3aBc + 3abC):1abc - комплементарное взаимодействие генов; 48:9:6:1 - 48(27АВС + 9АВс + 9АbС + 3Аbс):9аВС:6(3aBc + 3abC):1abc - эпистаз, эпистатор - аллель А и т. п.
Важно помнить, что на небольших выборках достаточно трудно или даже невозможно различить некоторые расщепления, связанные с разным числом генов, например 3:1 и 13:3; 1:2:1 и 9:3:4; 9:7 и 37:27 и т. п. Статистическая проверка по %2 не позволяет, как уже указывалось в главе III, отдать предпочтение одной из них, если по χ2 не отвергаются обе гипотезы. В таких случаях обычно повторяют опыт, увеличивая выборку, или (иногда) выбор между гипотезами делают на основе формальной логики. Очевидно, что во всех случаях необходимо делать генетическую проверку всех гипотез.
В реципрокных скрещиваниях кроликов с шерстью нормальной длины и длинношерстными (ангорскими) в первом поколении все крольчата имели шерсть нормальной длины, а в F2 среди 105 потомков 57 имели нормальную, 27 длинную и 21 - очень короткую шерсть. Скрещивание гибридов F1 с длинношерстными родителями дало расщепление - 39 длинношерстных, 15 короткошерстных и 18 нормальных. При скрещивании гибридов F1 с короткошерстными из F2 (индивидуальные скрещивания) давало разные результаты: с одними - расщепление в соотношении -3 нормальных : 3 короткошерстных : 2 длинношерстных; с другими - расщепление 1 нормальный : 1 короткошерстный. Объяснить расщепления, определить число генов, по которым различаются исходные формы, и их генотип.
Единообразие в F1 реципрокных скрещиваний указывает на отсутствие сцепления признака с полом и гомозиготность исходных форм. Расщепление в F2 на три фенотипических класса нельзя объяснить действием одной аллельной пары, несмотря на то, что по методу %2 гипотеза о моногенном расщеплении и неполном доминировании не отвергается - соотношение фенотипов в F2 соответствует расщеплению 2:1:1 (χ2 = 2,51, p>0,30). Она отвергается логически уже по результатам F1: при неполном доминировании гибриды первого поколения должны иметь промежуточный характер проявления признака, в опыте же наблюдается полное сходство с одним из родителей. Поэтому предполагаем расщепление по двум генам. Тогда величина одного сочетания гамет равна 105:16 = 6,56, расщепление в опыте - 57 : 6,56= =8,7 (~9), 21 : 6,56=3,2 (~3), 27:6,56=4,1 (~4). Н0 - расщепление по двум независимо наследуемым генам в соотношении 9:3:4; χ2 = 0,19, p>0,90. Отклонение в опыте случайно, гипотеза не отвергается. Гены локализованы в аутосомах и наследуются независимо, взаимодействуя по типу комплементарности: АВ - нормальная шерсть, Аb (или аВ) - короткая шерсть, аВ (или Аb) и ab - длинная шерсть. Генетическая проверка - скрещивания гибридов F1 с длинношерстными (1) и короткошерстными (2) потомками из F2 также не отвергают предложенной гипотезы:

χ2 = 0,75, p>0,70. Отклонение случайно, гипотеза не отвергается. Данное скрещивание является анализирующим. Если бы расщепление шло по одному гену, то в этом скрещивании могло появиться только два фенотических класса в соотношении 1:1.
2. Короткошерстные особи F2, согласно нашей гипотезе, могут иметь либо генотип AAbb, либо генотип Aabb. Поэтому возможны два варианта скрещивания с короткошерстными потомками F2
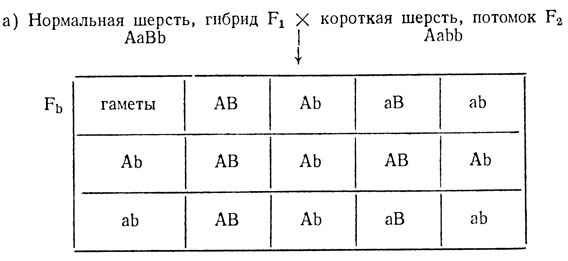
3АВ - нормальная шерсть, 3Аb - короткая шерсть, 2(аВ и ab) - длинная шерсть.

- Исходные формы различаются по двум комплементарным независимо наследуемым генам (9:3:4).
2. Генотипы исходных форм - 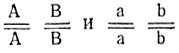 ; генотип
; генотип 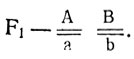
Скрестив реципрокно кур с белым и черным оперением, в первом поколении получили только белых цыплят, а во втором - 56 белых и 14 черных. Повторив опыт, во втором поколении получили 163 белых и 34 черных цыпленка. Объясните полученные данные. Чем можно объяснить разницу между первым и вторым опытом, если в них использовали одних и тех же кур и петухов? Каковы генотипы исходных птиц? Какую генетическую проверку можно провести для окончательного решения вопроса, о различиях между исходными формами?
Из результатов F1 можно предположить, что исходные формы гомозиготны и признак не сцеплен с полом. Расщепление в первом опыте дает возможность выдвинуть гипотезу о моногенном характере различий между родителями и расщеплении 3:1. Проверка по χ2 не отвергает данной гипотезы: χ2 = 0,93, p>0,70. Генотипы исходных форм: белая АА, черная аа. Проверка по χ2 той же гипотезы для второго опыта отвергает гипотезу о моногенном различии: χ2 = 6,29, p<0,05. Расщепление 163:34 можно объяснить, если принять, что родительские особи различаются по двум генам и расщепление 163:34 есть расщепление 13:3, χ2 = 0,28, p>0,80. (Расщепление в первом опыте - 56:14 - также может быть объяснено как 13:3, χ2 = 0,07, p>0,70.)
Гены исследуются независимо и взаимодействуют по типу доминантного эпистаза: IA, Iа и ia - белая окраска, iA - черная окраска, генотип гибридов Fi-IiAa, генотипы исходных птиц IIаа - белые, iiАА - черные.
Рассмотренный пример наглядно демонстрирует значение величины выборки - большие выборки увеличивают разрешающую способность генетического анализа при установлении числа генов в расщеплении.
Очевидно, что для окончательного вывода необходимо провести генетическую проверку обеих гипотез, например поставить анализирующее скрещивание, для чего полезно иметь в коллекции анализатор, в данном случае белых птиц с генотипом iiаа.
IV.1.2. Расщепления при локализации одного гена в Х-хромосоме, других - в аутосомах
При локализации одного из генов, контролирующих признак, в Х-хромосоме, в первом поколении реципрокных скрещиваний следует ожидать разных результатов: в одном проявится крисс-кросс наследование, в другом - единообразие. Расщепления в F2 прямого и обратного скрещиваний также должны быть разными, и характер расщеплений зависит от того, какой пол является гетерогаметным, а также от числа и типа взаимодействия генов. Основную формулу расщепления можно найти по теории вероятностей. При гетерогаметности мужского пола (например, у дрозофилы) и различию по двум генам, один из которых в Х-хромосоме, формулы расщепления определяются следующим образом.
Пусть ген А локализован в аутосоме, а ген В - в Х-хромосоме. Тогда в реципрокных скрещиваниях должны получиться oследующие результаты.

Общее расщепление по двум генам (записываются фенотипические радикалы):

Это основные формулы расщеплений в реципрокных скрещиваниях при взаимодействии двух генов с указанной локализацией, когда разные их сочетания (АВ, Аи, аВ и ab) имеют собственное фенотипическое проявление.
При других типах неаллельных взаимодействий какие-либо сочетания генов могут оказаться фенотипически неразличимыми, что изменяет характер расщеплений. Например, 3:5-3АВ:5(3Аb + 1аВ + 1аb) в одном, 9:7 - в другом направлении реципрокных скрещиваний; 7:1 - 7(3AB + 3Ab + 1ab):1аВ и 13:3 и т. д. Их легко определить, используя теорию вероятностей и зная расщепления по каждому гену.
Полезно запомнить, что если в Х-хромосоме исходной особи гомогаметного пола локализован доминантный аллель, то расщепление по изучаемому признаку (без учета пола) совпадает с расщеплениями при аутосомной локализации двух генов в соответствии с типом их взаимодействия. Однако некоторые фенотипические классы будут представлены только особями гетерогаметного пола. У потомков гомогаметного пола расщепление происходит по аутосомному гену (или оно вообще отсутствует); среди потомков гетерогаметного пола оно идет в соответствии с формулой 3:3:1:1 (или ее производной). В обратном скрещивании, где в Х-хромосоме особей гомогаметного пола локализован рецессивный аллель, расщепление в F2 происходит одинаково у потомков обоих полов.
Чтобы убедиться в этом, можно самостоятельно вывести формулы расщеплений в реципрокных скрещиваниях для разных типов неаллельных взаимодействий - комплементарного, эпистаза (с геном-эпистатором в аутосоме или в Х-хромосоме), полимерии.
При скрещивании самки дрозофилы с ярко-красными глазами с самцом, имевшим коричневые глаза, в первом поколении получили самок с красными, а самцов - с ярко-красными глазами. В обратном скрещивании все самки и самцы первого поколения имели красные глаза. Расщепления в F2 обоих скрещиваний приведены ниже.
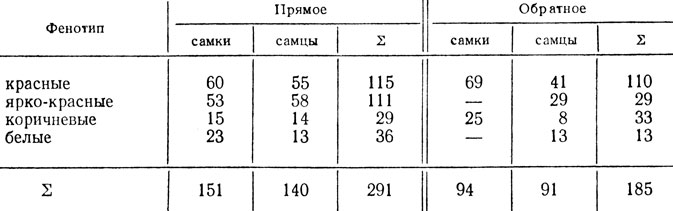
Объяснить расщепления и определить генотипы исходных мух и гибридов первого поколения.
Поскольку в F1 в одном из направлений реципрокных скрещиваний у самцов проявляется признак матери, а у самок - признак дикого типа (красные глаза), можно сделать два предположения: признак, по-видимому, сцеплен с полом; проявление у гетерозиготных самок F1красных глаз при реципрокных скрещиваниях мутантных форм свидетельствует о том, что мутации не-аллельны и исходные формы различаются не по одному гену.
Появление в F2четырех фенотипических классов также указывает на участие в расщеплении не менее двух генов. Гены не сцеплены, так как в расщеплениях нет преобладания родительских фенотипов. Нулевая гипотеза (Н0) - исходные формы различаются по двум взаимодействующим генам, один из которых локализован в Х-хромосоме, поэтому анализ расщеплений проводится в прямом и обратном скрещиваниях отдельно.
- Анализ расщепления в прямом скрещивании. Расщепление среди самцов и самок одинаково - можно предполагать, что исходная самка была рецессивна по гену, локализованному в Х-хромосоме. Тогда общая формула расщепления 3:3:1:1 и величина одного сочетания гамет - 291:8 = 36,4, расщепление в опыте - 111:36,4 = 3,0, 115:36,4 = 3,3; 30:36,4 = 0,8; 35:36,4 = 0,96, т. е. примерно 3:3:1:1; χ2 = 1,52, p>0,50. Отклонение в опыте случайно, гипотеза не отвергается.
- Анализ расщепления в обратном скрещивании. Н0 - та же, что и при анализе прямого скрещивания, но ожидаемое расщепление 9:3:3: 1, χ2 = 1,48, p>0,50. Отклонение случайно, гипотеза не отвергается. У самок расщепление по одному аутосомному гену в соотношении 3:1, χ2 = 0,10, p>0,70. У самцов расщепление по двум генам в соотношении 3:3:1:1, χ2 = 3,38, p>0,30.
Исходные формы различаются по двум генам, один из которых локализован в Х-хромосоме (обозначим его А), другой - в аутосоме (В). Гены наследуются независимо и взаимодействуют по типу комплементарности: АВ - красная окраска, Аb (или аВ) - коричневая, аВ (или Аb) - ярко-красная, ab - белая.

При скрещивании самки дрозофилы с красными глазами с самцом, имевшим ярко-красные глаза, в F1 все мухи имели красные глаза, в F2 произошло расщепление (а).
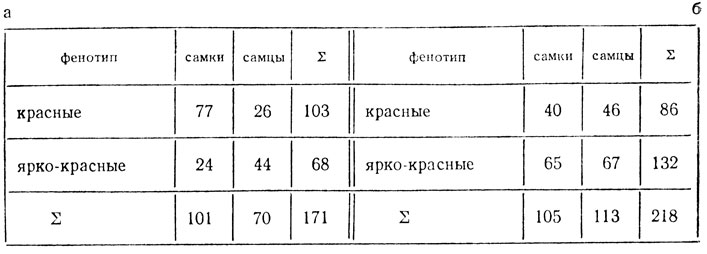
В F1 обратного скрещивания самки имели красные глаза, самцы - ярко-красные, в F2 произошло расщепление (б). Объяснить расщепления, определить генотипы исходных мух.
Из F1 реципрокных скрещиваний следует, что признак сцеплен с полом, так как в одном из них крисс-кросс наследование.
Рассмотрим результаты F2 отдельно в каждом скрещивании.
1. Расщепление в прямом скрещивании явно отличается от моногенного и оно разное у самок и самцов. Можно предположить, что родительские формы различаются по двум генам, один из которых в Х-хромосоме, причем самка маркирована доминантным аллелем этого гена (например, А). Тогда величина одного сочетания гамет (по сумме особей обоих полов) - 171:16 = 10,7, расщепление в опыте - 103:10,7 = 9,6; 68:10,7 = 6,4 Н0 - расщепление по двум генам 9:7, наследование независимое, χ2 = 1,07, p>0,30. Отклонение в опыте случайно, гипотеза не отвергается. Гибридные самки F1 гетерозиготны по обоим генам, их генотип 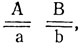 генотип самцов
генотип самцов 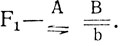 . Исходя из этого, а можно записать генотипы исходных мух.
. Исходя из этого, а можно записать генотипы исходных мух.
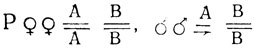
- красные глаза; 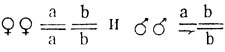 - ярко-красные глаза.
- ярко-красные глаза.
Среди самок расщепление произошло только по аутосомному гену (В) 3:1, χ2 = 0,08, p>0,80. Отклонение в опыте случайно, гипотеза не отвергается. Среди самцов расщепление идет по двум генам, величина одного сочетания гамет 70:8 = 8,75, расщепление в опыте 26:8,75 = 2,97 и 44:8,75 = 5,0 (~3 красных : 5 ярко-красных); χ2 = 0,003, p>0,95. Отклонение случайно, гипотеза не отвергается.
2. В обратном скрещивании в F2 расщепление у особей обоих полов одинаковое - преобладают мухи с ярко-красными глазами. Очевидно, исходная самка была рецессивна по гену, локализованному в Х-хромосоме. Гипотеза та же, что и в прямом скрещивании, расщепление 3:5, χ2 = 0,35, p>0,50.

Исходные мухи различаются по двум независимо наследуемым генам, один из которых локализован в Х-хромосоме (А), другой в аутосоме (В). Гены взаимодействуют по типу комплементарности, причем каждый из них так же как и двойной рецессив, вызывает развитие ярко-красной окраски глаз - Аb, аВ, ab; красная окраска проявляется только при наличии в генотипе доминантных аллелей обоих генов - АВ.
При расшифровке расщеплений удобно использовать формулы, по которым определяют число типов гамет, число фенотипов и возможных сочетаний гамет. Например, если расщепление происходит по трем генам, число возможных гамет и фенотипов равно 23 = 8. Правило выписывания гамет основано на законе чистоты гамет, согласно которому в каждую гамету попадает один аллель от каждой аллельной пары. Следовательно, среди 8 типов гамет, образуемых тригибридом Аа Bb Cc DD, четыре типа будут содержать аллель А, четыре - аллель а:
По гену В также четыре из 8 гамет получат аллель В, четыре - аллель b, которые могут попасть в гамету либо с А, либо с а:
Аллели Сие также будут представлены в гаметах с равной вероятностью в сочетании с аллелями генов А и В:
По гену D расщепления не происходит, следовательно, аллель D должен быть во всех гаметах. Таким образом, гибрид AaBbCcDD может образовать 8 типов гамет, которые соответствуют возможным фенотипам в расщеплении (если нет неаллельных взаимодействий, при которых некоторые классы соединяются в один):
Если гены наследуются независимо, частоты гамет разного типа равновероятны. Если гены сцеплены, частоты разных типов гамет определяются частотой рекомбинации между генами.
Вероятность выщепления каждого (любого) фенотипа в F2 и Fa определяют по теории вероятностей. При независимом наследовании генов вероятность проявления любого их сочетания в фенотипе определяется как произведение вероятностей выщепления каждого из них. В F2 вероятность появления доминантного фенотипа по каждому гену равна 3/4, рецессивного - 1/4. В Faвероятность выщепления любого фенотипа одинакова - (1/2)n. Например, появление потомков с фенотипом ABC в потомстве двух тригетерозигот - 3/4 (А)×3/4 (В)×3/4 (С) = 27/64, с фенотипом АВс - 3/4 (А)×3/4 (В)×1/4 (с) = 9/64, с фенотипом Abc - 3/4 (A)×1/4 (b)×1/4 (с) = 3/64 и т. д. Нетрудно заметить, что коэффициент для каждого фенотипа, в котором проявляется один доминантный аллель, всегда 3 - 3Аbс, 3аВс, 3аbС; коэффициент фенотипа, в котором проявляются два доминантных аллеля разных генов, 9 - 9АВс, 9аВС, 9АbС; три - 27 - АВС и т. д. Аналогично можно найти вероятность выщепления особей с любым генотипом. Например, вероятность появления особей с генотипом AABBccDD в нашем примере равна 1/4 (АА)×1/4 (ВВ)×1/4 (cc)×1(D) = 1/64, AaBBccDD - 2/4 (Aa)×1/4 (BB)×1/4 (cc)×1(D) = 2/64, AaBbCCDD = 4/64; AaBbCcDD = 8/64 и т. д.
Коэффициент каждого гомозиготного по любому числу генов генотипа всегда 1: ААВВСС, ААВВсс, ааВВсс и т. д. Коэффициент гетерозиготных генотипов определяется числом генов, находящихся в гетерозиготном состоянии: это 2, если в гетерозиготном состоянии один ген (АаВВсс, ааВbСС и т. д.); 4, если в гетерозиготном состоянии два гена (AabbCc, ааВЬСс и т. д.); 8, если три гена - АаВЬСс. Воспользуемся этими правилами при решении следующей задачи.
Ниже приведены результаты реципрокных скрещиваний белоглазых особей дрозофилы из двух линий (№ 11 и № 17), поставленных для выяснения их идентичности.
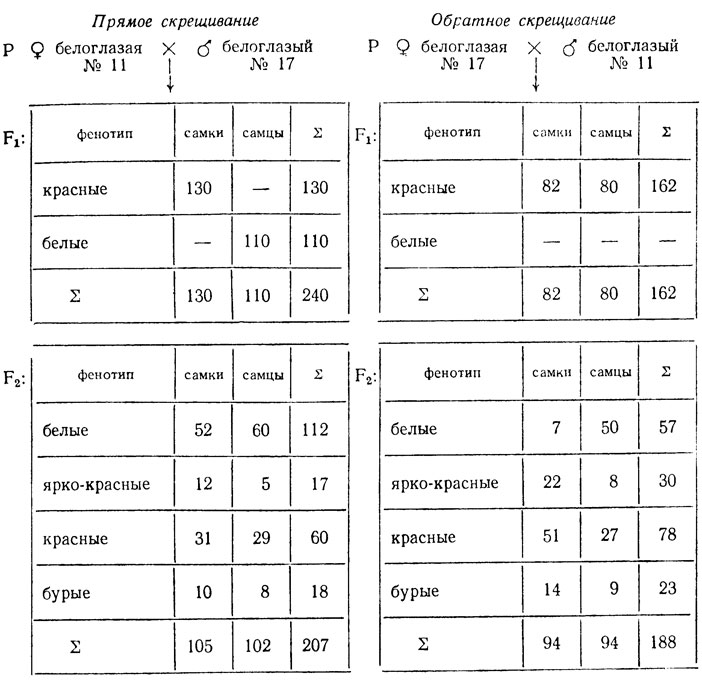
Объяснить расщепления, определить генотипы мух исходных линий и гибридов Fi.
Из F1 реципрокных скрещиваний можно предположить, что признак сцеплен с полом, так как в прямом скрещивании признак матери передается сыновьям. Появление в F1 гетерозиготных красноглазых самок (дикий тип) говорит о том, что мутации неаллельны и линии различаются не менее чем по двум генам, причем по крайней мере один из них локализован в Х-хромосоме. В связи с этим анализ F2 проводят отдельно по каждому скрещиванию.
1. Появление в F2 четырех фенотипических классов возможно при различии исходных форм по крайней мере по двум генам, один из которых локализован в Х-хромосоме (см. F1) (H0). Поскольку в прямом скрещивании наблюдается передача признака матери сыновьям, можно предположить рецессивность исходной самки из линии № 11 по гену, локализованному в Х-хромосоме. Тогда расщепление среди самок и самцов по этим генам должно быть одинаковым - 3:3:1:1; проверка по χ2 отвергает эту гипотезу - χ2 = 24,3, p<0,01. Расщепление на четыре стенотипических класса может быть следствием различия исходных форм и по трем независимо наследуемым генам, один из которых в Х-хромосоме. По каждому гену, локализованному в аутосоме, число возможных сочетаний гамет равно четырем (41), по гену, сцепленному с полом,- двум (21). Следовательно, возможно проявление 32 сочетаний гамет (если все гены наследуются независимо): 4×4×2 = 32.
Величина одного сочетания - 207:32 = 6,5, расщепление в опыте 112:6,5 = 17,2; 17:6,5 = 2,6; 60:6,5 = 9,2; 18:6,5 = 2,8 (≈17:3:3:9). Такое расщепление очевидно свидетельствует о взаимодействии этих генов, причем несколько классов неразличимы фенотипически.
Проверяем гипотезу, предположив, что белые составляют в расщеплении 17/32, красные - 9/32, бурые и ярко-красные по 3/32. Проверка по χ2 не отвергает этой гипотезы: χ2 = 0,48, p>0,90. Для определения генотипов исходных мух, а также для выяснения, какие взаимодействия трех генов приводят к образованию окрасок, полученных в опыте, вернемся к анализу F1. Самки F1 должны быть гетерозиготными по всем трем генам, самцы - гетерозиготны по аутосомным генам, а в их Х-хромосоме должен находиться рецессивный аллель.
Пусть в Х-хромосоме локализован ген А, в аутосомах - В и С. Тогда генотип особей F1 можно записать так: 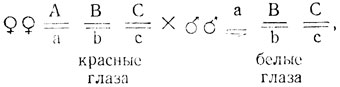 из чего следует, что сочетание генов ABC обусловливает развитие красной окраски глаз, аВС - белой.
из чего следует, что сочетание генов ABC обусловливает развитие красной окраски глаз, аВС - белой.
В F2 по теории вероятностей в этом случае возможно появление следующих классов: (1А:1а)×(3B:1b)×(3С:1с) = 9АВС:3АВс:3АbС:1Аbс:9аВС:3аВс:3аbС:1abc. Два класса в расщеплении определяются сразу по результатам F1; красные глаза (АВС) и белые глаза (аВС). Запишем эти окраски против соответствующих фенотипических радикалов в расщеплении. Поскольку при наличии в генотипе рецессивного аллеля а, локализованного в Х-хромосоме, и двух других генов (В и С) мухи имеют белые глаза, можно предположить, что аллель а является рецессивным эпистатором и тогда все особи с этим аллелем окажутся белоглазыми (aBc, abC и abc); мухи с ярко-красными и бурыми глазами составляют в опыте по 3/32. Эти окраски зависят от генов В и С, локализованных в аутосоме, если в Х-хромосоме есть аллель А. Например, АВс - ярко-красные, АbС - бурые (или наоборот). Тогда сочетание Abc должно давать белую окраску глаз, так как в опыте доля белоглазых составляет 17/32 - 9aBC+3aBc+3abC+1abc+1Abc = 17.
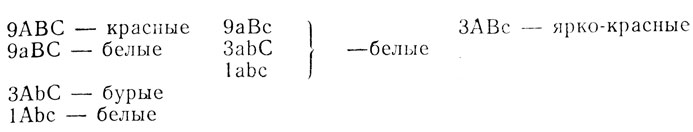
2. В F1 обратного скрещивания самки и самцы красноглазые.
Их генотипы: 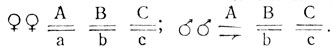
В F2обратного скрещивания также возникло четыре фенотипических класса, причем расщепления среди самок и самцов разные - среди самок преобладают мухи с красными, среди самцов - с белыми глазами. Очевидно, что появление четырех фенотипических классов у самок свидетельствует о том, что у них произошло расщепление по двум генам, локализованным в аутосомах. Ожидаемое при этом расщепление при независимом наследовании двух комплементарно взаимодействующих генов должно быть 9:3:3:1. Проверка по χ2 не отвергает этой гипотезы: χ2 = 0,11, p>0,95. Расщепление среди самцов должно идти по формуле, которая установлена при анализе прямого скрещивания: 17 белые:9 красные:3 ярко-красные:3 бурые. Проверка по χ2 не отвергает этой гипотезы: χ2 = 2,08, p>0,50.
Общее расщепление по трем генам среди особей обоих полов определяют, записав возможные фенотипические радикалы, их соотношение и окраску глаз на основе проведенного анализа.

Следовательно, расщепление должно соответствовать формуле: 27 красных:9 бурых:9 ярко-красных:19 белых. Проверка по %2 не отвергает данной гипотезы: χ2 = 1,02, p>0,70.
В результате анализа установлено, что фенотипически одинаковые линии генетически не идентичны. Они различаются по трем независимо наследуемым генам, один из которых (А) локализован в Х-хромосоме. Гены, локализованные в аутосомах (В и С), взаимодействуют по типу комплементарности. Рецессивный аллель гена, локализованного в Х-хромосоме, эпистатичен в отношении аутосомных генов.
Генотипы исходных линий
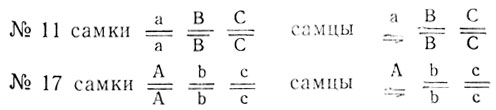
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'