3.1.4. X-сцепленные типы наследования
У человека каждый брак можно рассматривать как менделевский бэккросс (возвратное скрещивание) в отношении X- и Y-xpoмосомы:

Это означает, что в среднем мужская и женская зиготы формируются в отношении 1:1. Однако в действительности это не совсем так. Соотношение полов при рождении (известное как вторичное соотношение полов в отличие от первичного соотношения полов при оплодотворении) немного сдвинуто в сторону мальчиков (102-106 мальчиков на 100 девочек). Первичное соотношение полов точно неизвестно, но имеются некоторые данные, что оно также изменчиво. (Опубликовано множество работ по изучению первичного и вторичного соотношения полов. Результаты хромосомных исследований материала абортов, теоретически отражающие первичное соотношение полов, указывают на вероятное наличие равного соотношения мужских и женских эмбрионов. Однако как первичное, так и вторичное соотношение полов зависит от продолжительности периода между половым актом и овуляцией, от частоты актов, общих условий, включая даже состояние войны и мира. При искусственном оплодотворении доля мужских потомков оказывается существенно выше [704].) Формальные характеристики X-сцепленных типов наследования легко выводятся из механизмов определения пола.
X-сцепленный рецессивный тип наследования. Если использовать символ А для обозначения доминантного аллеля нормального дикого типа и символ а для рецессивного аллеля, то возможны следующие браки:
а) AA♀ × A♂. Все дети будут иметь фенотип A. Как этот, так и аналогичный ему брак aa × a неинформативны для генетического анализа.
б) AA♀ × a♂. У всех сыновей присутствует один из нормальных материнских аллелей, все они здоровы. Все дочери - гетерозиготы Aa. Они фенотипически здоровы, но являются носительницами аномального аллеля. В аналогичном браке aa♀ × A♂ Все сыновья поражены (а) и все дочери гетерозиготны (Aa).
в) Aa♀ × A♂. Этот тип брака наиболее важен для генетического анализа. Все дочери фенотипически нормальны, и половина из них являются гетерозиготными носительницами. Аналогичный брак Aa♀ × a♂ встречается очень редко. В этом случае ожидается отношение 1:1 пораженных и гетерозигот среди дочерей и отношение 1:1 пораженных и здоровых среди сыновей.
Основные формальные характеристики X-сцепленного рецессивного наследования следующие. Обычно пораженными являются мужчины, а для редких X-сцепленных заболеваний это справедливо почти всегда. Все их фенотипически здоровые дочери являются гетерозиготными носительницами. Среди сыновей гетерозиготных матерей соотношение пораженных и непораженных 1:1.
Строго говоря, передача признака от пораженных дедов через здоровых матерей пораженным внукам не может служить вполне убедительным доказательством локализации гена в X-хромосоме. Аналогичные рассуждения справедливы и в случае аутосомного гена, проявление которого ограничено мужским полом. Решающим является тот факт, что все сыновья пораженных мужчин здоровы. Однако этот критерий не подходит, когда заболевание настолько тяжелое, что больные не могут иметь детей.
Двумя наиболее известными и с практической точки зрения важными примерами являются две формы гемофилии, А и В (30670, 30690). На рис. 3.13 показана знаменитая родословная потомков королевы Виктории в европейских королевских домах. Одним из гемофиликов был царевич Алексей в России. По-видимому, власть Распутина над императорской четой основывалась, по крайней мере частично, на его способности успокаивать царевича. Были описаны родословные и большего размера, вероятно, наиболее обширная из них - с гемофилией В из Тенна (Швейцария). Однако на практике родословные, как правило, много меньше. Часто имеется только одно сибство с пораженными братьями, или вообще больной оказывается единственным пораженным в семье. Как и при доминантных заболеваниях (разд. 3.1.2), это вызвано сниженной репродуктивной способностью пораженных, что ведет к элиминации большого количества генов гемофилии в одном или в нескольких последующих поколениях после возникновения новой мутации. Как и следует ожидать, большинство больных гемофилией - мужчины. Однако имеется несколько исключений. На рис. 3.14 показана родословная (из Чехословакии), в которой гемофилик женился на гетерозиготной женщине, приходившейся ему двойной двоюродной сестрой, поскольку в поколении их родителей двое братьев женились на сестрах. Обе гомозиготные сестры имели гемофилию средней степени тяжести подобно их пораженным родственникам-мужчинам.
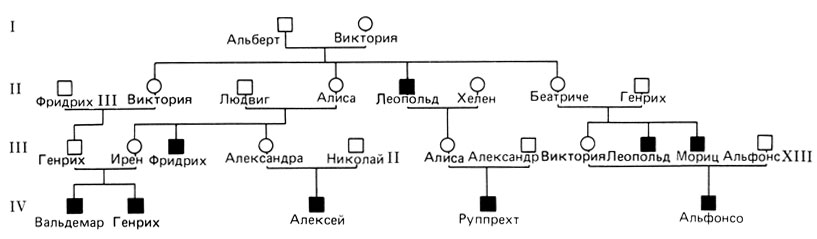
Рис. 3.13. Родословная с X-сцепленной рецессивной гемофилией А в европейских королевских домах. Королева Виктория (1.2) была гетерозиготой. Она передала мутантный ген одному сыну-гемофилику и трем дочерям
![Рис. 3.14. Родословная с двумя женщинами, гомозиготными по X-сцепленной гемофилии. Родители - двойные двоюродные сибсы. O - облигатные гетерозиготы [840]](pic/000140.jpg)
Рис. 3.14. Родословная с двумя женщинами, гомозиготными по X-сцепленной гемофилии. Родители - двойные двоюродные сибсы.
 - облигатные гетерозиготы [840]
- облигатные гетерозиготы [840]Другое X-сцепленное рецессивное заболевание-это синдром Леша-Найхана (30800) - редкая аномалия метаболизма пуринов, связанная с недостаточностью фермента HGPRT (гипоксантин-гуанин-фосфорибозилтрансфераза), которая приводит к тяжелой гиперурикемии, неврологическим расстройствам и неукротимому стремлению к самоповреждениям (рис. 3.15). Это заболевание послужило моделью для решения многих биохимических и генетических проблем, в частности, была найдена селективная система для идентификации и изоляции in vitro мутантных клеток с этим дефектом (разд. 4.2.2.6).
![Рис. 3.15. Родословная с синдромом Леша-Найхана [203]](pic/000142.jpg)
Рис. 3.15. Родословная с синдромом Леша-Найхана [203]
Некоторые X-сцепленные заболевания характеризуются значительной распространенностью. Наиболее часто встречаются дефекты цветового зрения, полиморфные варианты фермента глюкозо-6-фосфат - дегидрогеназы (G6PD) (разд. 4.2), а также X-сцепленная задержка умственного развития с маркерной (ломкой) X-хромосомой (разд. 8.2.1.2).
X-сцепленный доминантный тип наследования. X-сцепленное доминантное заболевание проявляется у гемизиготных мужчин и гетерозиготных женщин. Однако все сыновья пораженных отцов и здоровых матерей не несут патологических признаков, здоровы и их дети. С другой стороны, все дочери пораженных отцов поражены. Среди детей пораженных матерей должно наблюдаться тоже сегрегационное отношение 1:1 независимо от пола ребенка, так же как и при аутосомно-доминантном типе наследования. Если пораженные индивиды имеют нормальную репродуктивную способность, то в популяции больные женщины встречаются примерно в два раза чаще, чем больные мужчины. Поскольку лишь дети пораженных отцов дают возможность различить X-сцепленное доминантное и аутосомно-доминантное наследование, в случае малочисленных семейных данных трудно или даже невозможно сделать однозначный вывод о характере наследования.
Первый четкий пример X-сцепленного доминантного типа наследования был описан Сименсом (1925) [871]. Это кожное заболевание (30880), при котором образуется фолликулярный гиперкератоз, что в свою очередь приводит к частичной или полной потере ресниц, бровей и волос на голове (рис. 3.16). Однако тяжелые формы этого заболевания в данной родословной ограничены только мужчинами.
![Рис. 3.16. Родословная с фолликулярным кератозом. Первый пример X-сцепленного доминантного наследования у человека. ■ - тяжелая форма, ○ - легкое проявление, ⊗ - умерли до начала заболевания [817]](pic/000143.jpg)
Рис. 3.16. Родословная с фолликулярным кератозом. Первый пример X-сцепленного доминантного наследования у человека. ■ - тяжелая форма, ○ - легкое проявление, ⊗ - умерли до начала заболевания [817]
С тех пор для всех признаков с установленным X-сцепленным доминантным наследованием было подтверждено, что в среднем мужчины поражены тяжелее, чем женщины. В этом нет ничего неожиданного, поскольку у гетерозиготных женщин частичная компенсация может определяться нормальным аллелем. Полностью этот факт стал объясним после открытия феномена случайной инактивации одной из X-хромосом у женщин (разд. 2.2.3.3).
Другим примером X-сцепленного доминантного наследования служит устойчивый к витамину D рахит с гипофосфатемией (30780) [958]. В родословной на рис. 3.17 все 11 дочерей пораженных отцов страдали рахитом или гипофосфатемией, тогда как все десять их сыновей были здоровы. Пораженные матери имели как пораженных, так и здоровых сыновей и дочерей. Вероятность того, что такое же распределение (т. е. у пораженных отцов дочери всегда поражены, а сыновья всегда здоровы) может быть получено при аутосомно-доминантном наследовании, меньше чем 1:10000. Кроме того, в этой семье, как правило, мужчины больны тяжелее женщин.
![Рис. 3.17. Родословная с Х-сцепленным доминантным рахитом, устойчивым к витамину D, и гипофосфатемией. ■ - гипофосфатемия с рахитом, - гипофосфатемия без рахита. [958]](pic/000144.jpg)
Рис. 3.17. Родословная с Х-сцепленным доминантным рахитом, устойчивым к витамину D, и гипофосфатемией. ■ - гипофосфатемия с рахитом,
 - гипофосфатемия без рахита. [958]
- гипофосфатемия без рахита. [958]X-сцепленное доминантное наследование при летальности мужчин-гемизигот. Как уже отмечалось, у женщин X-хромосомные заболевания обычно имеют менее тяжелые проявления, чем у мужчин. В некоторых случаях поражение мужских зигот оказывается настолько сильным, что они погибают еще до рождения. Тогда в родословных среди пораженных должны быть только женщины, а среди их пораженных детей - только дочери, причем в соотношении со здоровыми дочерьми и сыновьями 1:1:1. Кроме того, мужские гемизиготы, которые не погибают на очень ранней стадии беременности, должны обнаруживаться в спонтанных абортах или среди мертворожденных мальчиков. Ленц (1961) [759] первым показал, что этот тип наследования существует у человека для заболевания, известного под названием incontinentia pigmenti (пигментный дерматоз) (Bloch-Sulzberger) (30830).
В перинатальном периоде у девочек, пораженных этим заболеванием, развиваются воспалительный эритематоз и везикулярные кожные поражения. Позже появляется "мраморная" пигментация (рис. 3.18, А). Синдром иногда сочетается с аномалиями зубов. На рис. 3.18, Б показана типичная родословная. Важно, что аутосомно-доминантный тип наследования с проявлением, ограниченным женским полом, также может рассматриваться как альтернативная гипотеза. Обе эти гипотезы предсказывают следующие соотношения.
![Рис. 3.18. А. Больная с синдромом Блоха - Сульцбергера (пигментный дерматоз) (Fuhrmann, 1974 [433]); отчетливо виден мраморный рисунок кожи. Б. Родословная с пигментным дерматозом, · - спонтанный аборт; ● - пигментный дерматоз [759]](pic/000145.jpg)
Рис. 3.18. А. Больная с синдромом Блоха - Сульцбергера (пигментный дерматоз) (Fuhrmann, 1974 [433]); отчетливо виден мраморный рисунок кожи. Б. Родословная с пигментным дерматозом, · - спонтанный аборт; ● - пигментный дерматоз [759]
а) При аутосомно-доминантном ограниченном полом наследовании (после соответствующей корректировки, разд. 3.3.4) для пораженных и здоровых сестер пробандов ожидается отношение 1:1. Все братья будут здоровы. Если предположить, что в популяции соотношение полов 1:1, то среди здоровых сибсов следует ожидать соотношение полов 2♂:1♀. С другой стороны, при X-сцепленном наследовании ожидаемое количество здоровых братьев намного меньше, поскольку предполагается, что половина мужских зигот погибает до рождения (что, вероятно, ведет к повышению уровня спонтанных абортов). Среди здоровых сибсов ожидается отношение 1♂:1♀.
б) При аутосомно-доминантном наследовании аномальный ген может наследоваться как от отца, так и от матери. Следовательно, и среди более отдаленных родственников как по отцовской, так и по материнской линии можно ожидать пораженных. С другой стороны, при X-сцепленном наследовании ген должен передаваться от матери. В случае очень редкого заболевания среди родственников со стороны отца патологии быть не должно.
в) При аутосомно-доминантном наследовании утрата мутантных генов (в расчете на поколение) должна быть относительно небольшой по сравнению с общим числом этих мутаций в популяции, поскольку здоровые мужчины-носители должны обладать нормальной репродуктивной способностью. Следовательно, при генетическом равновесии (разд. 5.1.3.1) лишь малая доля случаев заболевания в популяции будет связана с новыми мутациями. С другой стороны, при X-сцепленном наследовании утрата зигот должна быть высокой вследствие гибели гемизигот. Следовательно, многие случаи заболевания в популяции должны объясняться новой мутацией, т. е. обширные родословные будут редкими [924].
Имеющиеся статистические данные подтверждают гипотезу об X-сцепленном доминантном наследовании при летальности мужских гемизигот. Кэрни (1976) [613] сообщает о 693 случаях среди женщин и 16 случаях среди мужчин. При этом у 55,4% больных женщин имело место семейное отягощение. Как можно объяснить спорадические случаи у мужчин? Конечно, хорошо известно явление "проход через огонь" (Durchbrenners). Хэдрон [696] для обозначения случайно выживших индивидов с летальным генотипом использовал термин "беглецы" (escapers), но Ленц (1975) [760], основываясь на гипотезе Гартлера и Франка (1975) [676], предложил более оригинальное объяснение. По его мнению, мутация затрагивает только одну из цепей двойной спирали ДНК (либо в сперматозоиде, либо в ооците).
К этой же группе можно отнести и некоторые другие заболевания. Одно из них проявляется некоторыми уродствами рта и языка, медианной заячьей губой и синдактилией особого типа (31120) [674]. Другими примерами могут служить очаговая гипоплазия кожи, X-сцепленная пятнистая хондродисплазия, недостаточность орнитинтранскарбамилазы (31125; летальная у новорожденных гемизиготных мальчиков) и частичная липодистрофия с липотрофным диабетом. Судя по опубликованным родословным, такой тип наследования возможен и, например, в случае специального варианта мышечной дистрофии плечевого пояса [565], [943].
Гены, локализованные в Y-хромосоме. До 50-х гг. большинство генетиков были убеждены в том, что Y-хромосома у человека содержит гены, которые изредка мутируют, давая Y-сцепленный (или голандрический) тип наследования с передачей признака от отца сыновьям, т. е. пораженными оказываются только мужчины. Однако Штерн в своем обзоре [897] приводит ряд доказательств того, что освященный временем и ставший учебным пример Y-сцепленного наследования тяжелой формы ихтиоза у человека уже не может считаться убедительным. Единственный признак, для которого Y-сцепленное наследование все еще обсуждается, - это волосатые ушные раковины, т. е. наличие волос на внешнем крае уха. Было опубликовано несколько обширных родословных, которые демонстрировали передачу признака от отца сыновьям. Однако поздний возраст проявления (преимущественно в третьем десятилетии жизни), а также крайне изменчивая экспрессивность и высокая популяционная частота (до 30% в некоторых популяциях) весьма затрудняют дискриминацию голандрического наследования от мультифакториального с ограничением по полу. Иначе говоря, для этого признака однозначно принять Y-сцепленное наследование не представляется возможным.
Эйхвальд и др. (1955) [643] описали детерминированный Y-хромосомой трансплантационный антиген у мыши, который они назвали HY. Авторы предположили, что он является одним из факторов половой дифференцировки, в частности, мужских гонад. В соответствии с гипотезой, высказанной Оно [1248], HY-антиген экспрессируется во всех клетках мужского организма, но только гонадные клетки имеют HY-рецептор, связывающий этот антиген. Связанная с рецептором молекула активирует развитие тестикулярной ткани. Для проверки этой важной для понимания механизмов детерминации пола гипотезы были предприняты обширные исследования [1342] (см. разд. 4.7.5).
AHTH-HY-антисыворотка мыши вступает в реакцию с клетками всех тестированных до сих пор млекопитающих, включая человека. Даже клетки птиц и амфибий дают перекрестную реакцию. Следовательно, этот антиген в ходе эволюции остается почти неизменным.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'