3.4.1. Классические подходы в экспериментальной генетике: эксперименты по скрещиванию и гигантские хромосомы
Согласно третьему закону Менделя, сегрегация двух разных пар аллелей происходит независимо друг от друга; все возможные зиготы по двум парам аллелей формируются при свободной рекомбинации. При скрещивании гетерозиготы AaBb и гомозиготы aabb образуются в равных пропорциях четыре типа особей.
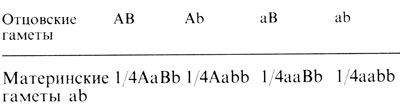
Вскоре после переоткрытия законов Менделя Бэтсон, Сандерс и Пеннет (1908) [561] нашли исключение из этого правила у Lathyrus odoratus. Одни комбинации встречались чаще, а другие - реже, чем ожидалось. В некоторых случаях в потомстве чаще встречались родительские типы (в нашем примере AB - отцовское растение, а ab - материнское), в других случаях - два других типа Ab и aB.

θ - частота рекомбинантов
Создавалось впечатление, что у каждого из родителей аллельные гены либо притягиваются, либо отталкиваются. Бэтсон и др. [561] предложили для первого случая термин "притяжение", для второго - "отталкивание". Морган (1910) [448] указал, что притяжение и отталкивание отражают расположение двух генов на одной или на гомологичных хромосомах. Он ввел термин "сцепление". Притяжение означает, что у дважды гетерозиготного родителя гены A и B расположены на одной хромосоме AB/ab отталкивание означает, что они расположены на гомологичных хромосомах Ab/aB. Для обозначения положения генов в фазах притяжения и отталкивания чаще употребляются термины цис и транс соответственно. При полном сцеплении потомство может быть только двух типов. Однако в большинстве случаев обнаруживаются все четыре типа, хотя два из них - в меньшем количестве. Морган объяснил это явление обменом хромосомными участками между гомологичными хромосомами во время мейотического кроссинговера. Он обнаружил также, что частота кроссинговера зависит от расстояния между локусами двух генов на хромосоме. Используя в качестве аналитического инструмента рекомбинационный анализ, Морган и его коллеги успешно локализовали большое количество генов у дрозофилы. Их результаты были подтверждены, когда в начале 30-х гг. Гейтц, Бауэр и Пэйнтер открыли гигантские хромосомы у некоторых двукрылых и сопоставили данные о локализации конкретных генов, полученные косвенными методами, со структурными перестройками определенных хромосом. С тех пор анализ сцепления проведен для огромного количества видов.
Сцепление и ассоциация. Иногда предполагают, что сцепленные гены в популяции должны ассоциировать, т. е. хромосомные комбинации AB и ab (притяжение) должны обнаруживаться чаще, чем комбинации Ab и aB (отталкивание). Однако для популяции со случайным скрещиванием это не так. Даже при тесном сцеплении повторяющийся во многих поколениях кроссинговер будет приводить к равномерному распределению в популяции всех четырех комбинаций AB, ab, Ab, aB. Как правило, ассоциация генетических признаков не указывает на сцепление, а вызвана другими причинами.
Однако это правило имеет исключения. Некоторые комбинации тесно сцепленных генов на самом деле встречаются чаще, чем ожидается при равномерном распределении. Такое "неравновесие по сцеплению" впервые было постулировано у человека для групп крови Rh (разд. 3.5.4) и доказано для главного комплекса гистосовместимости (МНС), особенно для системы HLA (разд. 3.5.5), а также для ДНК-полиморфизмов. Неравновесие по сцеплению имеет две причины.
- Исследуемая популяция образовалась из двух популяций, различающихся частотами аллелей A, a и B, b, а время, прошедшее с момента смешения, недостаточно для полной рандомизации.
- Высокая частота определенных аллельных комбинаций сцепленных генов поддерживается естественным отбором.
Детальнее эти вопросы будут обсуждаться в связи с системой МНС (разд. 3.5.5) и при обсуждении ассоциации между HLA и разными заболеваниями (разд. 3.7.3).
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://genetiku.ru/ 'Генетика'